Impact Factor
- Issue 14; 2026
- Issue 13; 2026
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Volume 16; 2026
- Advance Articles
- Past Issues
- Cover Images
- Cover Suggestion
- Index & Coverage
- Special Issues
1. Introduction
2. Mechanosensitive Transducers...
3. Gene Delivery Systems
4. Optimization Strategies for...
5. Visualization
6. Applications of Sonogenetics
7. Conclusion and Future Outlook
Abbreviations
Acknowledgements
References
International Journal of Biological Sciences
International Journal of Medical Sciences
 Global reach, higher impact
Global reach, higher impact

Theranostics 2026; 16(13):7345-7366. doi:10.7150/thno.134803 This issue Cite
Review
Sonogenetics for precision medicine: from molecular toolkit to clinical translation
Xinrui Yu1, Min Pu1, Ruoli Wang2, Haitao Ran1 ![]()
1. Department of Ultrasound, the Second Affiliated Hospital of Chongqing Medical University, Chongqing, 400010, China.
2. Department of Neurosurgery, The Second Affiliated Hospital, Chongqing Medical University, Chongqing, China.
Received 2026-3-20; Accepted 2026-5-26; Published 2026-6-4
Abstract

Sonogenetics is an emerging technology for precise biological modulation. It utilizes ultrasound waves for remote, noninvasive, and spatiotemporally precise functional intervention in target cells genetically engineered for acoustic sensitivity. This review systematically summarizes the three core components comprising sonogenetics: sound-responsive agents for modulating cellular functions, highly efficient vectors for targeted gene delivery, and ultrasound emitters with optimized parameters. Furthermore, we delineate optimization strategies for these components and propose a standardized workflow for sonogenetic operations. This review proposes strategies to accelerate the transformation process of sonogenetics by analyzing current limitations in regulatory efficiency and clinical safety. Finally, this review discusses the existing technical challenges and transformation bottlenecks, and points out the promising directions of future research.
Keywords: sonogenetics, mechanosensitive ion channels, targeted gene delivery, microbubbles, precision medicine
1. Introduction
Sonogenetics is a promising biomedical regulatory strategy that can be used to remotely control molecular and cellular activities in living organisms [1]. This technology integrates ultrasound and genetic engineering, which can accurately regulate target cells. It can not only overcome the inherent depth limitations of optogenetics but also solve the problem of insufficient spatial resolution unique to systemic pharmacology. Sonogenetics shows immense promise in noninvasive treatment of diseases and the exploration of untouchable anatomical areas ranging from deep brain neural circuits to visceral tumors.
According to their different physical mechanisms, sonogenetics can be divided into two categories [2-4] (Figure 1): mechanical sonogenetics and thermal sonogenetics. Mechanical sonogenetics mainly utilizes acoustic radiation forces (ARF) and cavitation effect of ultrasound [5]. These mechanical forces directly trigger the conformational changes of the exogenously expressed mechanosensitive (MS) ion channels in target cells, thus regulating the excitability of cells [6]. Thermal sonogenetics uses the thermal energy generated by ultrasound [7]. This local temperature increase activates thermoresponsive elements, such as heat-shock promoters or thermoresponsive repressors, regulating gene expression [8]. Although thermal sonogenetics strategies have practical value, their activation mechanisms and core design principles are different from those induced by mechanical effects. This review focuses on the strategies of mechanical sonogenetics.
Classification of sonogenetics. Mechanical sonogenetics utilizes ARF and cavitation. Ultrasound directly triggers conformational changes of the exogenously expressed MS ion channels in target cells, producing a series of biological effects. Thermal sonogenetics uses thermal energy generated by ultrasound to activate thermoresponsive elements, such as heat-shock promoters or thermoresponsive repressors, to regulate gene expression. Created with BioRender.com. ARF: acoustic radiation force; MS: mechanosensitive.
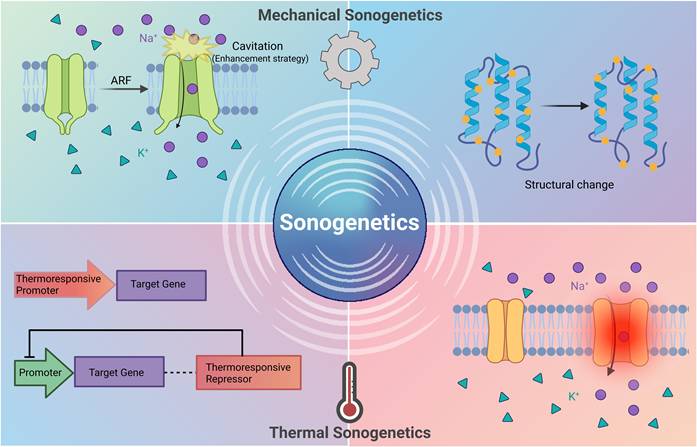
The concept of "sonogenetics" was first proposed in 2015. With the help of microbubbles, researchers successfully activated the neurons that expressed TRP-4 ion channel of Caenorhabditis elegans by low-frequency ultrasound [9]. This pioneering work not only lays the theoretical foundations for sonogenetics, but also demonstrates the potential of combining ultrasound with mechanosensitive media to accurately regulate the function of biomolecules. Since then, this field has entered a period of rapid development, and the scope of research has expanded from the activation of basic nerves in simple model organisms [9] to the regulation of complex physiological functions in mammalian models [10]. The success of these studies shows that sonogenetics has developed into a noninvasive biological regulation tool with high spatiotemporal precision. A complete sonogenetics system usually includes the following tools: (1) an ultrasonic transmitter used to provide precise ultrasonic stimulation [11]; (2) a sonosensitive mediator as the core of the technology; (3) an efficient and safe gene vector [12]; (4) microbubbles to amplify ultrasonic mechanical forces [13,14], enhance gene delivery efficiency [15], open the blood-brain barrier (BBB) [16] and assist imaging [17], (5) reporter genes to track the expression site and efficiency of the target gene [18].
Although sonogenetics has made rapid progress, it still faces major obstacles to the translation from laboratory research to clinical technology. This review systematically outlines the core toolkit of mechanical sonogenetics and puts forward a clear optimization path. On this basis, we proposed a comprehensive technological workflow covering all aspects from imaging guidance to precise treatment. At the same time, this paper lists the main application areas of sonogenetics at present. This review compares sonogenetics with other precise modulation techniques, such as optogenetics and chemogenetics, in multiple dimensions to provide researchers with fresh research perspectives. Furthermore, this paper assesses the primary bottlenecks encountered in the clinical transformation of sonogenetics and proposes targeted solutions. With its continuous development, sonogenetics is expected to become a safe, efficient, and multifunctional precise treatment technology in clinical practice.
2. Mechanosensitive Transducers in Sonogenetics
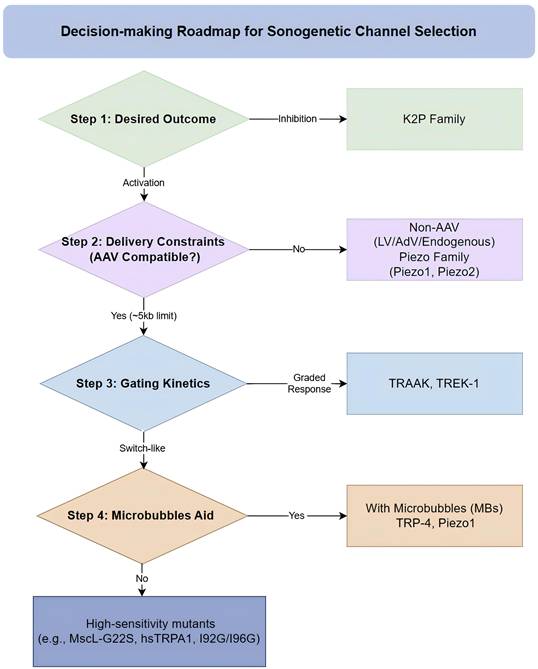
Mechanosensitive transducers are the mediators that convert ultrasound-induced mechanical stimuli into electrochemical signals (Table 1). The activation of these mechanosensitive transducers depends on two biophysical mechanisms: the bilayer model and the tether model [19,20]. MS ion channels are common sonogenetic mediators. The ideal candidate is considered to have the characteristics of high acoustic sensitivity and expression without disrupting basal cellular physiology. By continuously optimizing mechanosensitive proteins, scientists have achieved a paradigm shift from microbubble-assisted activation to direct channel activation. To facilitate the selection of appropriate sonogenetic tools, we propose a strategic roadmap (Figure 2). The roadmap matches candidate channels with specific experimental goals. However, research results on activation thresholds, desensitization rates, and ion selectivity are often reported under different experimental conditions, and there is still a lack of a unified standardized comparison framework. The establishment of evaluation indicators is critical to promote sonogenetics from laboratory to clinical practice.
Summary of research on mechanosensitive channels in sonogenetics.
| Family | Mutations | Subjects (in vitro/in vivo) | Ion channel type | Ultrasonic response/pressure | Safety | Ref. |
|---|---|---|---|---|---|---|
| MscL | MscL-G22S | ·HEK 293T cells and mouse primary neurons ·Neurons in the cerebral cortex or dorsomedial striatum of mice | Non-selective large-pore (>30 Å) ion channel, allowing passage of ions (including Ca2+) and small proteins. | 500 kHz center frequency, 1 kHz PRF, 40% duty cycle | Expression of MscL-G22S shows no obvious side effects judged by mouse behavior, body weight, and resting membrane potential of neurons. | [12] |
| MscL-I92G/I96G | ·A549 cells ·mouse | Non-selective pore (30 Å in diameter) that allows easy passage of ions and small molecules below 1 kDa | 0.044 MPa sound pressure (in vitro) 0.053 MPa sound pressure, equivalent to ISPTA of 15.2 W/cm2 (in vivo) | / | [28] | |
| MscL I92L | ·Rat hippocampal neurons in primary culture | Slightly more permeable to cations than anions | 0.25 MPa peak negative pressure | / | [27] | |
| TRP | TRP-4 | ·ASH, AWC, PVD sensory neurons and AIY interneurons of C. elegans | A stretch-sensitive, pore-forming mechanosensitive cation channel | 2.25 MHz, 10-ms pulse, peak negative pressures of 0.41, 0.47, and 0.6 MPa (require gas-filled microbubbles) | Use low ultrasound peak negative pressures to avoid inertial cavitation. | [9] |
| TRPP1/2 TRPC1 | ·Primary murine cortical neurons (endogenous expression) | Calcium-selective mechanosensitive ion channels | 300 kHz, 15 W/cm2, 500 ms (CW) | No sustained calcium accumulation or irreversible membrane perforation was found after repetitive stimuli. | [35] | |
| hsTRPA1 | ·HEK 293T cells ·Mouse layer V motor cortical neurons (in vivo) | A widely conserved calcium-permeable nonselective cation channel | 7 MHz transducer; 2.5 MPa (in vitro), 0.35-1.05 MPa (in vivo); 100 ms duration | hsTRPA1-expressing neurons could be repeatedly stimulated without apparent deleterious effects on cell health or a substantial decrement in calcium flux. | [36,69] | |
| TRPC6 | ·Cultured mouse cortical neurons and hippocampal neurons (endogenous expression) | A mechanosensitive nonselective cation channel | 1 MHz, 190 mW/cm², 500 ms | / | [37] | |
| Piezo | Piezo1 | ·HEK293T cells, Jurkat T cells, PBMCs | Nonselective cation channels with a modest preference for Ca2+ over Na+ | 2 MHz transducer, ∼0.6 MPa acoustic pressure (require gas-filled microbubbles) | Does not cause damage to target cells. Ultrasound induces minimal alteration to target cells. | [44] |
| Prestin | Prestin (N7T, N308S) | ·SH-SY5Y cells ·C57BL/6J mice | Anion transporters | 0.5 MHz, 0.5 MPa, 0.1% duty cycle, 3 s transient | Gene expression decreased markedly within 7 days in vivo. | [51] |
| Prestin (N548S, V715G) | ·HEK293T cells, HeLa cells, Primary neuron ·Hippocampal neurons and motor cortex of C57BL/6JNarl (B6) mice | Anion transporters | 0.5 MHz, 0.5 MPa, 10 Hz PRF, 2000 cycles per burst, 3 s duration | No tissue disruption or hemorrhage are observed. | [52] | |
| K2P | TRAAK | ·Xenopus laevis oocytes | Mechanosensitive K+ channels | 10 ms, 5 MHz, 1.2/3.6 W/cm2 | The activation does not require cavitation or thermal effects, supporting the safety. | [61] |
| ASICs | ASIC1a | ·CHO, Primary cortical neurons ·Cortex, hippocampus (CA1/CA2, dentate gyrus) and amygdala of C57BL/6J mice, Asic1⁻/⁻ mice, Asic3⁻/⁻ mice | A sodium channel, resulting in the intracellular calcium elevation possibly by activating voltage-gated calcium channels | 1 MHz transducer; 7.4 mW/cm2 (ISPTA) at 700 mVpp (in vitro); 5 mW/cm2 (ISPTA) at 900 mVpp (in vivo) | Use ultrasound exposure below 10 mW/cm2 (ISATA) to ensure safe therapeutic applicability. | [63] |
| YAP | YAP | ·Murine C2C12 mesenchymal precursors (endogenous expression) | Mechanosensitive transcriptional coactivator | 3.6 MHz excitation frequency, 100 Hz PRF, 27.8% duty cycle and ISATA = 44.5 mW/cm2 | / | [65] |
| MEC | MEC-4 and MEC-6 | ·C. elegans (endogenous expression) | DEG/ENaC family mechanosensitive channels | SAW: 27.4 MHz frequency, 3 MPa acoustic pressure, 5 W electronic power. | / | [68] |
CHO: the Chinese hamster ovary; PBMCs: peripheral blood mononuclear cells; PRF: pulse repetition frequency; ISPTA: spatial peak temporal average intensity; ISATA: spatial average temporal average intensity; SAW: Surface acoustic wave.
A roadmap for the selection of sonogenetic channels. This flowchart mainly divides the mechanosensitive transducers into four dimensions: desired outcome, delivery constraints, gating kinetics, and acoustic adjuvants. Based on this flowchart, researchers can make preliminary selections of mechanosensitive transducers according to the usage purpose and scenarios. AAV: adeno-associated virus; AdV: adenovirus; LV: lentivirus; MBs: microbubbles.
2.1 MscL in sonogenetics
The large-conductance mechanosensitive channel (MscL) was originally isolated from Escherichia coli. Its function is similar to an emergency release valve, which can protect bacteria from osmotic lysis [21]. Structurally, MscL is composed of 5 identical subunits with a large pore diameter (25–30 Å) [22], offering a high conductance pathway for ions and small molecules. MscL has two transmembrane helices (TM1 and TM2) and an extracellular loop connecting two transmembrane regions. As the main part that forms the "pore" of the channel, the TM1 helix can alter channel behavior, such as sensitivity to mechanical stimulation [22-24].
The concept of "sonogenetics" was proposed in 2015. Yet as early as 2014, Heureaux et al. conducted pioneering research consistent with the core principles of sonogenetics. Heureaux et al. developed a tetracycline-regulatable adenoviral system to efficiently introduce wild-type (WT) MscL and its gain-of-function mutant MscL G22S into mammalian retinal pigment epithelial (RPE) cells. However, this approach required microbubble-mediated acoustic tweezing cytometry (ATC) rather than direct ultrasound activation [25]. Subsequent studies have substantiated that some gain-of-function mutants could be activated by low-intensity focused ultrasound (LIFU) alone. The MscL-G22S mutant enabled region-selective neuromodulation in mouse brains via LIFU [12]. Recent studies comparing MscL-G22S, MscL-G22N, and MS ion channels under identical conditions have further demonstrated the superior ultrasonic sensitivity of MscL-G22S [26]. It has been demonstrated that the I92L mutant [27] and the I92G/I96G mutant are capable of lowering activation thresholds, with the latter exhibiting a significantly reduced activation threshold (0.044 MPa) compared to the I92L mutant and endogenous Piezo1 [28,29]. Although significant progress has been made in engineering MscL variants to optimize acoustic sensitivity, MscL's non-selective ion permeability poses risks of intracellular ion imbalance and cytotoxicity during long-term expression.
2.2 TRP in sonogenetics
Transient Receptor Potential (TRP) channels enable organisms to respond to various environmental stimuli. According to the homogeneity of amino acid sequences, the TRP channel superfamily is divided into seven subfamilies [30]. They are involved in a range of other physiological and pathological conditions. As cationic channels, most members of the TRP superfamily can be activated by various stimuli. In addition to being mechanosensitive, TRP can respond to voltage, temperature, and ligands [31,32].
Initially, TRP-4 was identified as an essential pore-forming subunit of native mechanotransduction channels [33]. Subsequently, ultrasound was demonstrated to affect TRP-4 MS ion channels expressed in C. elegans with the help of microbubbles to influence animal behavior [9,34]. In mammalian systems, several TRP channels have been identified as responsive to ultrasound. Specifically, TRPP1/2 and TRPC1 are activated by ultrasound, leading to the influx of extracellular Ca²⁺, while the influence of cavitation and temperature has been ruled out [35]. Moreover, the TRPM4 ion channel has been shown to act as a signal amplifier, enhancing the calcium accumulation signal triggered by the opening of MS channels [35]. In the search for optimal sonogenetic tools, the human Transient Receptor Potential A1 (hsTRPA1) channel was demonstrated to exhibit superior responsiveness to ultrasound stimuli at 1 MHz, 2 MHz, and 7 MHz compared to other candidates such as MscL, Prestin, TRPV1, and Piezo1. The ultrasound sensitivity of hsTRPA1 likely depends on the structure of its N-terminal tip region and its interactions with the actin cytoskeleton and cholesterol [36]. These findings provide a foundation for the subsequent design of mutants to improve sonogenetic efficiency. In endogenous contexts, TRPC6 has been proposed to act as a biosensor for ultrasound-induced mechanical distortions in the lipid bilayer of the mammalian brain. The activation is accompanied by sodium influx, resulting in the excitation of neuronal networks [37]. While endogenous channels such as TRPP1/2, TRPC1, and TRPC6 enable "gene-free" modulation, they carry a risk of non-specific activation (off-target effects) in non-targeted tissues exposed to ultrasound. Furthermore, ultrasound neuromodulation via endogenous MS channels does not constitute true sonogenetics, as it lacks the genetic manipulation necessary for the precision of sonogenetics.
2.3 Piezo in sonogenetics
The Piezo family, including Piezo1 and Piezo2, is evolutionarily conserved [38]. Piezo1 and Piezo2 possess a 38-transmembrane (TM) helix topology and form a homotrimeric propeller-shaped structure with a central ion-conducting pore and three peripheral mechanosensing blades. The curved TM region of three blades forms a nano-bowl configuration that can expand the in-plane membrane area, potentially conferring high mechanosensitivity to Piezo channels. [39,40]. Piezo1 is activated by externally applied poking, stretching, and shear stress, or endogenously originating local membrane tension and myosin II-mediated traction force. In contrast, while Piezo2 is effectively activated by poking, it responds relatively poorly to stretch [41]. Given the difference, Piezo1 is more widely used in sonogenetics.
The mechanosensitivity of Piezo1 has been harnessed for diverse therapeutic strategies. In cancer immunotherapy, ultrasound stimuli, assisted by microbubbles, activate endogenous Piezo1 in primary T cells (PBMCs) and Jurkat cells, resulting in a significant upregulation of anti-CD19 CAR (chimeric antigen receptor) gene expression [42]. To further enhance specificity and reduce energy requirements, Piezo1-targeted microbubbles (PTMBs) were developed [43]. Furthermore, in keloid treatment, 111 kHz low-frequency ultrasound was shown to inhibit fibroblast migration and increase total apoptosis of primary keloid fibroblasts (PKF) without the assistance of microbubbles by activating Piezo1 channels to trigger Ca²⁺ influx [44].
Because Piezo channels widely exist in multicellular organisms, future research may explore the differential expression of Piezo family channels across various tissues and their diverse functions within specific tissues when exposed to ultrasound. This is expected to further expand the application of ultrasonic regulation in biological functions and disease treatment. Moreover, as a typical large transmembrane protein [45], Piezo1 cannot be delivered efficiently through the mainstream AAV (adeno-associated virus) vectors. This limitation is mainly due to the fact that the size of its coding sequence exceeds the packaging capacity of AAV.
2.4 Prestin in sonogenetics
Prestin belongs to the SLC26 family of anion transporters and exhibits piezoelectric properties. It is different from the classical ion channels. It is a voltage-dependent anion transporter in mammalian cochlear outer hair cells. It converts changes in transmembrane voltage into cellular length alterations, thereby enabling high-frequency hearing and cochlear amplification [46-48]. Furthermore, Prestin has also been observed to respond to ultrasound via ultrasonic-driven conformational transitions [49], generating electrical motion and functional modulation in cells.
Engineered Prestin (N7T, N308S mutant) was first utilized by Huang et al. as a sonogenetic tool to modulate mammalian cells and neurons while eliminating interference from the auditory pathway [50]. Another study employed US-responsive DNA (Prestin plasmid, pPrestin)-loaded MBs (pPrestinMBs) to achieve noninvasive gene delivery by transiently disrupting the BBB, followed by ultrasound activation. Activation was observed in 58.2% of pPrestin-expressing cells, significantly higher than the activation rate in pPrestin-transfected cells without ultrasound activation [51]. Recently, the Prestin (N548S, V715G) variant, enhancing prestin oligomerization, was developed through unbiased screening to further increase the efficacy of sonogenetic neurostimulation [52]. These experiments suggested that ultrasound can effectively activate neurons expressing Prestin.
2.5 K2P in sonogenetics
Among the six two-pore domain potassium (K2P) subfamilies [53], only the TREK-1, TREK-2, and TRAAK channels within the TREK subfamily show mechanosensitivity [54]. Mechanical force is transmitted directly to TRAAK and TREK via the lipid bilayer. Additionally, they also exhibit polymodal gating properties, responding to various physical and chemical stimuli such as arachidonic acid, temperature, pH, and lipids [55-57]. Activation of mechanosensitive K+ channels leads to K+ efflux and subsequent hyperpolarization [58]. This channel family is significant to sonogenetics, as it represents the first and currently only inhibitory toolkit in sonogenetics.
Sorum et al. developed a quantitative description of TRAAK, TREK-1, and TREK-2 activation by membrane tension and excluded the influence of thermal and cavitation effects. Among these, TRAAK exhibits the highest sensitivity. Furthermore, the result demonstrated that low-intensity and low-frequency ultrasound can increase the membrane tension and thus activate TRAAK channels. Notably, the study only quantitatively analyzed the tension-response curve and did not verify the activation effect of ultrasound on TREK-1 and TREK-2. Their work revealed two different gating modes: TRAAK and TREK-1 can provide a graded K+ conductance proportional to the size of mechanical force, whereas the activation of TREK-2 occurs in a narrower range of force values. This "switch-like" gate is similar to MscL and Piezo1 [59-61]. Another study demonstrated at the molecular level that ultrasound increases the current flowing through TRAAK, TREK-1, and TREK-2 channels, providing direct evidence for their ultrasonic responsiveness [29].
Hence, these channels have wide applications in sonogenetics. TRAAK and TREK-1 are suitable for applications that require graded and proportional modulation, such as delicate neuronal activity, while TREK-2 may be utilized for threshold-dependent activation. Verifying these inhibitory tools in mammalian models is still the first task to achieve bidirectional sonogenetic control. However, because some K2P channels, such as TRAAK and TREK-1 [59,62], have endogenous expression, it is necessary to consider the off-target effects.
2.6 Potential candidates
In addition to currently known sonogenetic tools, the types of proteins that have been found to respond to ultrasound are still increasing. Recent studies have identified endogenous mechanotransducers with distinct ultrasound sensitivity, but their heterologous expression and application in sonogenetics remain underexplored. Investigating these candidates is significant for enhancing the precision and diversity of sonogenetic the toolkit.
In acid-sensing ion channels (ASICs), studies suggested that ASIC1a and ASIC3 can respond to ultrasound. ASIC1a is widely expressed in the brain. Lim et al. pointed out that ultrasonic stimuli can activate ASIC1a in the mouse brain through dynamic cytoskeletal rearrangement [63]. Similarly, ASIC3 has been shown to play a role in mediating therapeutic ultrasound on chronic muscle pain [64].
Yes-associated protein (YAP) is a transcriptional mechanotransducer, not an ion channel. Ultrasound stimulation can reduce the phosphorylation level of YAP at Serine 127, thus driving its accumulation in the nucleus, and ultimately promoting the expression of proliferation-related genes [65]. The study suggested that it has potential application value in regenerative medicine.
In touch receptor neurons, DEG/ENaC (degenerin/epithelial Na+ channel) protein plays a key role in mechanosensory transduction. Among these, the mec-4 gene is crucial to the mechanosensation of C. elegans [66]. MEC-6, a component of the degenerin channel complex, is involved in mechanotransduction within touch cells [67]. MEC-4 and MEC-6, expressed in the ALM and PLM neurons of C. elegans, have been demonstrated to jointly mediate neural activity and behavioral responses induced by ultrasound stimulation [68]. Additionally, both TRP-4 and MEC-4 cooperate to transduce ultrasound stimuli into behavioral changes in C. elegans [34].
3. Gene Delivery Systems
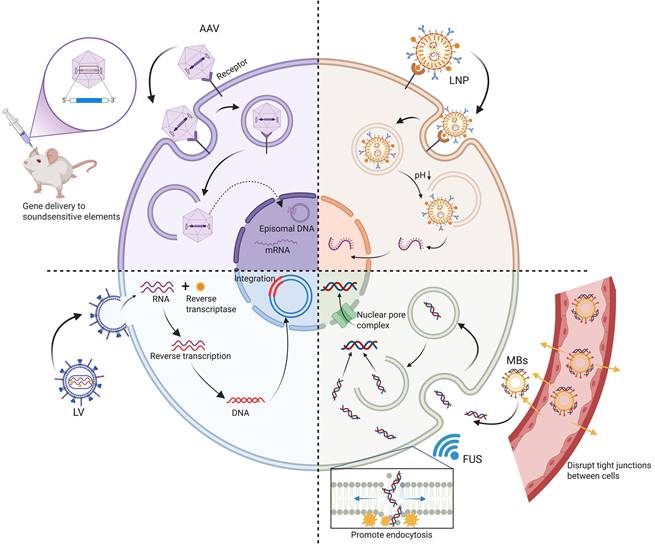
The efficient and safe delivery of MS channels is one of the key bottlenecks in the clinical translation of sonogenetics [70]. An ideal gene delivery system must not only ensure successful expression of MS ion channels in the target tissue, but also balance factors such as expression duration, transgene packaging capacity, tissue targeting specificity, immunogenicity, and delivery efficiency of the delivery vehicle. The current strategies are mainly divided into two modes: viral vector and non-viral vector (Figure 3). In terms of sonogenetics, these two delivery methods have their own advantages and limitations (Table 2).
The gene delivery system of sonogenetics. It illustrates viral (AAV, LV) and non-viral (LNP, FUS combined with MBs) vectors used to deliver mechanosensitive protein genes to target cells. AAVs enter cells via endocytosis and are stably expressed in a free form in the nucleus; LVs utilize reverse transcriptase to integrate genes into the host genome to achieve permanent expression; LNPs, as a non-viral platform, can protect and release mRNA into the cytoplasm; the combination of FUS and MBs can enhance local gene delivery in specific regions to reduce the off-target effect through sonopororation effect and other mechanisms. Created with BioRender.com. AAV: adeno-associated virus; FUS: focused ultrasound; LNP: lipid nanoparticle; LV: lentivirus; MBs: microbubbles.
Comparison of key characteristics, mechanisms, and properties of viral and non-viral gene delivery vectors.
| Core Components | Delivery mechanism | Gene capacity | Immunogenicity | Expression duration | Cell targeting | Ref. | |
|---|---|---|---|---|---|---|---|
| AAVs | Target gene expression cassettes + inverted terminal repeats (ITRs) + capsid proteins (VP1/VP2/VP3) | The AAV vector undergoes receptor-mediated endocytosis, achieves endosomal escape, and translocates to the nucleus to facilitate gene expression. | ~5 kb | Low, but faces the issue of pre-existing immunity against natural serotypes. | Transgene expression can endure for several years. | Serotypes naturally target different tissues. | [94-96] |
| LVs | Capsid proteins + unrelated virus + recombinant viral genome | Single-stranded RNA (ssRNA) is reverse-transcribed and semi-randomly integrated into the host genome. | ~9kb | Concerns regarding immunogenicity exist for in vivo applications. Innate immunity is easily triggered but controllable; adaptive immunity is low risk but requires vigilance. | Long-term stable expression is achieved due to the integration of the gene into host cells, but it carries the risk of insertional mutagenesis. | Alter the envelope glycoproteins (pseudotyping) to regulate tropism. | [97,98] |
| Adv | The protein capsid encapsulating the genome | Double-stranded DNA (dsDNA) is delivered and persists as free entities within the nuclei of transduced cells. | ~30kb | High | Medium-term expression | / | [99-101] |
| LNPs | Cationic or ionizable lipid, a helper lipid, a PEG-lipid, and cholesterol | Enter cells via endocytosis, then escape from the endosome. | Flexible | Significantly lower than viral vectors | Transient expression | Most intravenously administered nanoparticles accumulate in the liver. | [102,103] |
| MB-assisted US-guided gene delivery | Core (SF₆, C₃F₈, C₄F₁₀) + shell (polymers, proteins, surfactants, or phospholipids) | The gene is loaded into the MB shell layer. Ultrasound triggers the rupture of the MBs to release genes, synchronizing with the opening of the BBB. | Limited | Lower than viral vectors | Transient expression | Control of ultrasound irradiation area and modification of microbubbles | [104-106] |
AAV: adeno-associated virus; ITRs: inverted terminal repeats; VP: viral protein; LV: lentivirus; Adv: adenovirus; LNP: lipid nanoparticle; PEG: polyethylene glycol; MB: microbubble; US: ultrasound; BBB: blood-brain barrier.
3.1 Viral vectors
Viral vectors leverage the intrinsic infectivity of viruses to transduce host cells. They protect their internal genetic material from the degradation of host enzymes. Due to the high gene transduction efficiency of viral vectors, they are currently the primary delivery tools of sonogenetics.
As ~25 nm icosahedral viruses without an envelope [71], adeno-associated viruses (AAVs) are widely employed in sonogenetics research because of their inherent safety and high biocompatibility [72,73]. AAV genomes can achieve long-term transgene expression for several years as non-integrated episomes in the nucleus [74]. However, because the genes introduced by AAV are in a free state, the risk of their loss in the process of cell division is high [75]. Specific serotypes (e.g., AAV9) and engineered variants (e.g., AAV9.7m8) can accurately target the central nervous system (CNS) [76] and primary visual cortex (V1) [6]. The use of small-size cell-specific promoters like human synapsin (hSyn) or CaMKII further enhances specificity [6]. The delivery strategy has been verified in many sonogenetic studies, including the precise expression of MscL-G22S in primary neurons [12] and the expression of Cre-dependent TRPA1 in cortical neurons [36]. Despite these advantages, the application of AAVs is still subject to the inherent limitation of limited packaging capacity (~5 kb) [77]. This often forces researchers to strike a trade-off between functionality and specificity. This is a primary obstacle that hinders the wide application of AAVs in sonogenetics.
Lentiviral vectors (LVs) have larger packaging capacity (~9 kb) [78] and the ability to integrate into the genome. However, for in vivo applications, people are still concerned about insertional mutagenesis and immunogenicity. LVs have been employed to stably express MS channels such as TRPV1 [35] and MscL [28] in primary neurons and cancer models. The development of doxycycline-inducible (Tet-on) lentiviral system facilitates precise, temporally controllable expression of MscL [28]. Additionally, pseudotyping technology enhances the targeting efficiency of LVs. This is achieved by replacing envelope proteins to modulate the receptor-binding specificity [80]. Examples include using rabies virus glycoproteins to target neurons [81] or using VSV-G fused with anti-EGFR scFv pseudotype to target tumors [82].
Similar to AAVs, adenoviruses (AdVs) deliver genetic material without genomic integration [83]. AdVs can carry more than 30 kb of exogenous DNA, which is far more capacity than AAVs and LVs. However, their strong immunogenicity severely restricts their in vivo application [84]. Nevertheless, AdVs are still of great value in specific experimental situations. Heureaux et al. successfully utilized the recombinant AdV system to achieve high-efficiency, dose-dependent expression of MscL in retinal pigment epithelial cells [25].
3.2 Non-viral vectors
To avoid the inherent immunogenicity and production costs of virus vectors, the non-viral vectors have become an attractive alternative. Non-viral vectors will not be integrated into the genome of host cells, which eliminates the risk of insertion mutations and improves the safety of gene delivery. Moreover, these vectors can accommodate larger transgenes and are suitable for mass production. The two major challenges of non-viral gene delivery are the relatively low transfection efficiency and the typically transient expression. However, the short duration of transgenic expression is not an absolute disadvantage, depending on the treatment goal.
As a multifunctional, low-immunogenicity non-viral platform [85], lipid nanoparticles (LNPs) are used in sonogenetics [86]. By modifying their compositions, they can target specific cell types or change their metabolic processes. Their transient expression is suitable for short-term sonogenetic interventions, which helps to reduce the potential cytotoxicity caused by the long-term accumulation of exogenous MS channels. There has been a study using cationic nanoliposomes to deliver MscL I92L plasmids to tumor cells [87]. Despite these advantages, the in vivo application of lipid nanoparticles (LNPs) is still limited by their inherent liver accumulation effect and limited transfection efficiency.
Microbubble-assisted ultrasound-guided gene delivery is regarded as a promising gene delivery method, which is especially suitable for applications in the CNS [88]. Utilizing focused ultrasound (FUS) to trigger microbubble cavitation, this method can not only be used as an adjuvant to transiently overcome natural barriers, such as the BBB [89,90], but also directly act as a gene carrier [91]. This ensures that gene entry is strictly limited to the irradiated focal area, thus minimizing the off-target effect [92]. This approach has been successfully verified in the mouse brain. In the study, 1-MHz FUS and plasmid-loaded microbubbles were used to target the BBB and deliver the Prestin gene to specific neurons [51]. Despite its potential, the clinical translation necessitates the precise optimization of gene-loading capacity for large sequences [91], microbubble stability, and ultrasonic parameters [93].
4. Optimization Strategies for Sonogenetics
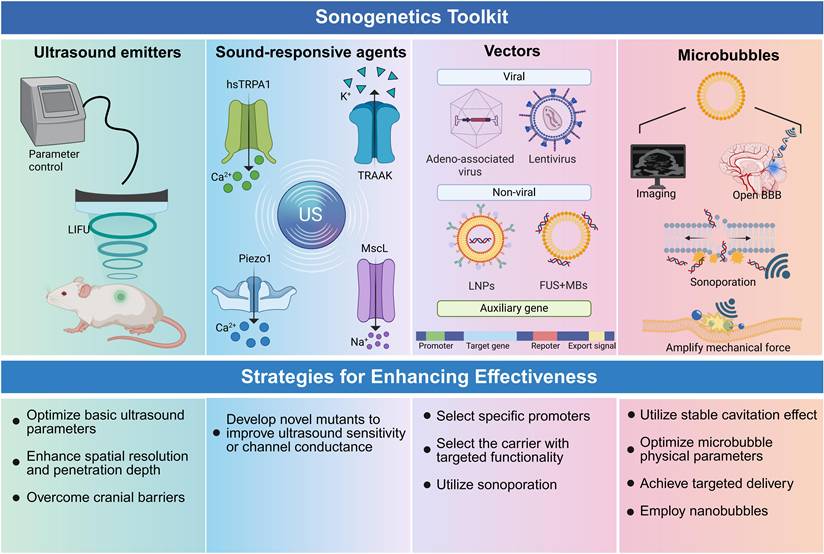
The clinical translation of sonogenetics hinges on enhancing its efficiency, safety, and precision. To achieve these objectives, we propose optimization strategies primarily focused on three dimensions: ultrasound stimulation parameters, mechanosensitive proteins, and microbubble adjuvants (Figure 4).
Sonogenetics toolkit and strategic pathways for enhancement. (1) Summarizes the essential components, including ultrasound emitters for parameter control, sound-responsive agents (e.g., hsTRPA1, Piezo1), viral and non-viral vectors for gene delivery, and microbubbles as amplifiers. (2) Outlines key optimization pathways to improve efficiency and safety regarding the aforementioned four aspects. Created with BioRender.com. BBB: blood-brain barrier; LIFU: low-intensity focused ultrasound; US: ultrasound.
4.1 Gas-filled microbubbles and nanobubbles
Gas-filled bubbles serve as amplifiers in sonogenetics by leveraging stable cavitation. These amplified forces facilitate the activation of MS ion channels in cell membranes at lower ultrasound energy. The modifiability of gas-filled bubbles allows for further system optimization. However, experiments need to avoid inertial cavitation that is induced by bubble rupture, resulting in tissue damage. Notably, if a system lacks genetic manipulation steps, regulation assisted solely by microbubbles does not constitute sonogenetics.
Microbubbles are the most commonly used sonoadjuvants. In C. elegans, MBs facilitate behavioral responses at low acoustic pressures (0.79 MPa). In contrast, the response differential between presence and absence of MBs diminishes at high pressures, as the high-pressure ultrasound alone provides enough mechanical force for activation. This confirms their role in lowering the energy barrier required for stimulation [34]. The amplification effect is size-dependent, with smaller MBs (1.5–2.5 μm) triggering stronger responses than larger variants (4–6.5 μm) [9]. Furthermore, PTMBs have been developed to enhance targeting specificity. This molecular targeting technology enables cell-specific activation and seven adhered MBs induce a stronger response than three [43]. Optimizing the number, size, and targeting ability of MBs is essential for advancing sonogenetic efficacy.
With the advancement in nanotechnology, gas-filled nanostructures have been used to enhance safety, precision, and efficacy of ultrasound-mediated neuromodulation. Hou et al. utilized gas-filled nanostructures, gas vesicles (GVs), to enhance ultrasound sensitivity. Under ultrasound pressure (1 MHz, 0.28 MPa), only harmonic signals were observed, indicating that GVs did not induce the collapse of gas bubbles to damage tissue. This result confirms that nanostructures have better safety [107]. Recently, PEGylated gas vesicles (PGVs) were developed with FDA-approved polymer shells to improve biocompatibility. These nanostructures facilitated the precise modulation of deep-brain circuits with extremely high spatial resolution. This method can distinguish brain regions only ~1.1 mm apart, and selectively induce different behaviors such as "rotation" and "freezing" [10]. Compared with microbubbles, nanobubbles are easier to penetrate the tissue gaps, achieving noninvasive targeted delivery and supporting high-precision spatial neuromodulation.
4.2 Ultrasonic equipment and ultrasound parameters
The ultrasound stimulation device controls the key acoustic parameters that affect sonogenetic results, including frequency, intensity, pulse duration, and waveform [108], by modulating mechanical forces, thermal effects, and cavitation effects [109]. Ideally, the stimulation device should provide flexible parameter optimization to adapt to the kinetic properties of target MS channels. Finding the balance is essential to improve activation efficiency and reduce risks, including desensitization, excessive thermal deposition, or inertial cavitation.
Currently, there is a lack of unified standards for ultrasound parameters in this field. Sound pressure is usually the direct driving force that turns on the MS channel [12]. The choice of frequency usually depends on the required depth of tissue penetration, which must be balanced with spatial precision. Deep tissue requires low-frequency ultrasound to ensure sufficient penetration [11]. In the study of visual restoration, the selection of frequency also needs to consider the repetition rates required for visual recovery [6].
Once channels are activated and achieve the expected effect, the focus should be turned to safety. When employing MBs, researchers should pay attention to the Mechanical Index (MI) to ensure that operation is carried out within a stable cavitation window [110]. The lower duty cycle (DC) emphasizes the mechanical effects of LIFU, which can improve the safety of sonogenetics. Intensity is defined as power per unit area. The total energy delivery is quantified through spatial peak pulse average intensity (ISPPA) and spatial peak temporal average intensity (ISPTA). The current ultrasound safety regulations are based on FDA diagnostic guidelines. It is stipulated that ISPPA should be less than 190 W/cm2, and ISPTA should be less than 720 mW/cm2, and MI should be less than 1.9 [111]. However, the safety parameters of therapeutic ultrasound still need to be further studied.
CNS regulation is an important application field of sonogenetics, and the development of related equipment can significantly enhance therapeutic efficacy. The cranium causes significant signal attenuation and distortion [112,113]. To solve this problem, time-shifting techniques within phased array transducers are used. They can control all waves to reach the target at the same time, so as to correct distortions [114]. For cortical regions, high-frequency ultrasound (15 MHz) can be transmitted through cranial windows implanted in mice to achieve precise stimulation of the V1 cortex [6]. In addition, a miniature wearable ultrasound system integrating a self-focusing transducer and bioadhesive hydrogel has been developed for long-term neuromodulation. This ultrasonic transmission can not only pass through the 8 mm human skull, but also achieve a smaller focal spot size and higher spatial resolution [115]. Equipment with high spatial resolution and portability is the key direction for the future development of sonogenetic hardware.
The collaborative integration of multiple enhancement strategies usually brings better results. Although LIFU has significant advantages in penetrating bones and tissues, its spatial resolution is inevitably affected by its longer wavelengths [11]. Combining low-frequency ultrasound with target-modified bubbles can theoretically resolve the trade-off. This combination may significantly enhance the interaction between ultrasound and tissue in the MS ion channel of the targeted cells by these target-modified bubbles, so that deep-tissue cells can be accurately activated with high spatial resolution. Such collaborative approaches are vital to give full play to the therapeutic potential of sonogenetics.
4.3 Ion channel mutants
MS ion channels are the core of sonogenetics. Some wild-type MS channels are naturally insensitive to LIFU, which limits the efficiency of sonogenetics. By engineering novel channel mutants with enhanced ultrasound sensitivity or conductance, safer, lower-pressure ultrasound can be used to elicit equivalent biological responses.
Mutants of the MscL channel represent the most thoroughly studied sonogenetic tools. The hydrophobicity of G22 determines pressure sensitivity of the MscL channel. The replacement of this residue with a hydrophilic residue can reduce the gating threshold [116]. The MscL-G22N mutant (Gly-to-Asn) damaged the hydrophobic "lock", reducing the pressure required for channel opening to below 5 mmHg [117]. In hypo-osmotic solution, the MscL-G22S mutant (Gly-to-Ser) expressed in retinal pigment epithelium (RPE) showed better activation effect than WT MscL [6]. The MscL-I92G/I96G double mutant was engineered to modify mechanosensitivity through TM1-TM2 interaction [118]. Its activation requires ultrasound pressure of only 0.044 MPa (26% Amplitude), which is significantly lower than the 0.053 MPa (31% Amplitude) required by WT MscL [28].
In addition to the mutants of MscL, analysis of Prestin protein sequence among non-echolocating and echolocating species revealed that the 7th and 308th amino acids often switch from N to T and from N to S, respectively. These observations led to the generation of Prestin mutants: Prestin (N7T), Prestin (N308S), and Prestin (N7T, N308S). The ultrasound sensitivity of mammalian cells expressing Prestin (N7T, N308S) was ~11-fold higher than that of control cells under low-frequency (0.5 MHz), low-energy (0.5 MPa and 0.1% of duty cycle), and transient (3 s) ultrasound conditions [50,51]. Future research should clarify the mechanisms of action of existing highly sonosensitive channels, explore novel mutant libraries to further optimize ultrasound responsiveness, and screen out new ultrasonic response candidate molecules from a variety of organisms.
5. Visualization
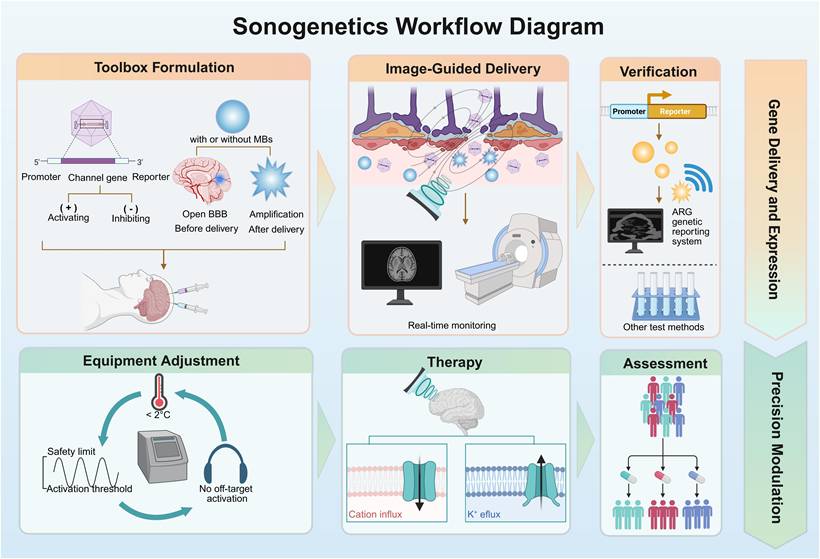
The lack of real-time visibility in the process of in vivo gene delivery poses significant safety concerns and hinders clinical translation. Visualization strategies aim to establish a closed-loop framework covering "precise delivery, real-time monitoring, and efficacy verification." This framework is designed to assess whether the genes have reached the target and been successfully expressed. This can meet the clinical demand for noninvasive and quantifiable treatment evaluation. Real-time imaging monitoring of gene delivery and reporter gene expression is the basis for providing safety and precision. Consequently, we propose a sonogenetic workflow that integrates visualization techniques, which makes sonogenetics have preliminary clinical feasibility (Figure 5).
Schematic diagram of the workflow of sonogenetics. The process is divided into two primary phases. The first stage is gene delivery and expression, including vector formulation, image-guided delivery, and verification; the second stage is precision modulation, which involves parameter optimization, therapeutic stimulation of MS channels, and efficacy assessment. Created with BioRender.com. ARG: acoustic reporter gene; BBB: blood-brain barrier; MBs: microbubbles; MS: mechanosensitive.
5.1 Image-guided gene delivery
Currently, imaging-guided gene delivery strategies have not been widely applied in basic sonogenetics. The delivery primarily relies on stereotactic or local injections to ensure that the gene vectors reach the target site directly. However, in view of the requirements of clinical translation for noninvasive precision, image-guided strategy has become a priority.
The following noninvasive monitoring methods hold promise for applications in sonogenetics. Magnetic resonance-guided focused ultrasound surgery (MRgFUS) is a pivotal technology: the three-dimensional imaging feature of MRI provides precise spatial localization for ultrasound focus, while FUS transiently opens the BBB to enhance vector concentration at the target site [119]. Human clinical trials delivering therapeutics have demonstrated there are no serious treatment-related side effects [120]. In nonhuman primates (NHPs) and humans, the technology has been used to facilitate the entry of AAV vectors carrying a green fluorescent protein (GFP) reporter gene and the hSyn promoter into the brain, yielding a 7–50-fold increase in GFP-positive neurons in the BBB-opened region compared to unopened regions [121]. Similarly, ultrasound has been used to guide the AAV9-GFP delivery during in utero fetal gene therapy of pigs. The turbulent flow in fetuses was observed to ensure precise amniotic cavity injection and avoid off-target effects [122]. Ultrasound and MB-guided adenoassociated viral vector (UMGAAV) utilizes microbubbles as real-time indicators of AAVs. The "blooming" artifacts induced by MB destruction confirm the treatment area, while increasing vector internalization [123].
5.2 Genetic reporting system
Reporter genes are introduced to indicate successful expression following delivery. These genes are co-delivered and co-expressed in the host cells together with the target gene. The expression of the reporter gene can reflect the activity of the target gene, the localization of the target protein, and the overall transduction efficiency.
Fluorescent reporter genes have become important tools for in vitro mechanism validation due to their intuitive visualization and high spatial and temporal resolution. After incident light that excites the wavelength reaches the fluorophore in the cell, the fluorescence will release photons of a distinct emission wavelength and be detected by a specialized camera [124,125]. Green fluorescent proteins (GFP), mCherry, and yellow fluorescent protein (YFP) are often used as reporters. In sonogenetic studies, GFP variants have been employed to visualize the expression of MscL-G22S in HEK293T cells [12] and Prestin constructs in primary neurons and brain slices [51]. Similarly, red fluorescent proteins like mCherry have been utilized to verify the successful expression of MscL mutants [28]. However, the application of fluorescent reporter genes in in vivo experiments is limited by their poor light penetration and background fluorescence interference. Other optical reporter genes also face similar problems. Hence, these reporter genes can only be used for surface tissue or in vitro analyses [126].
In contrast, acoustic reporter genes (ARGs) offer the advantages of deep tissue penetration, low background signal, and accurate localization, making them ideal for in vivo applications. ARGs are based on GVs. GVs are air-filled protein nanostructures found in certain waterborne bacteria and archaea. Their ability to scatter sound waves lays the foundation for them as reporter genes. In 2018, Bourdeau et al. first developed ARGs for noninvasive ultrasound imaging of intra-mammalian microbiomes based on genetically engineered GVs [127]. In 2019, ultrasound imaging of gene expression was successfully achieved in mammalian cells by engineering gene clusters to adapt to the transcription and translation systems of mammalian cells. Ultrasound images clearly revealed the localization of mARG-expressing cells. Calculating the signal difference between pre-collapse and post-collapse of GVs can eliminate background scattering from host tissues, which clearly confirms that the ultrasound signals originate from mARG-expressing cells [128]. Technical optimizations can further improve the imaging quality. The burst ultrasound reconstructed with signal templates (BURST) increased the sensitivity of ARGs ~1,000-fold using two pressure level ultrasound [129]. mARG_Ana, an improved mammalian ARGs, produces 38-fold stronger nonlinear contrast than first-generation versions. By applying the xAM pulse sequence, nonlinear signals are enhanced while linear background scattering is canceled out from host tissues. Unlike BURST, this method does not rupture GVs, enabling non-destructive imaging [130].
Notably, GVs are multifunctional, facilitating Theranostics for sonogenetics. Beyond imaging gene expression, GVs can also amplify mechanical forces through stable cavitation to enhance channel activation. Furthermore, the inertial cavitation effect induced by high-pressure ultrasound can produce therapeutic effects, such as targeted destruction of tumor tissues. These functions make ARGs a promising choice to replace optical reporters in noninvasive, deep-tissue imaging in sonogenetics.
6. Applications of Sonogenetics
Sonogenetics harnesses the unique synergy between the deep tissue penetration of ultrasound and gene-targeted cell specificity, and is emerging as a transformative noninvasive biomedical intervention. This section elaborates on the expanding transformation research prospects of sonogenetics, highlighting its core roles in three key areas, including the treatment of neurodegenerative and neuropsychiatric disorders through precise regulation of neural circuits, noninvasive restoration of sensory function, and the induction of anti-tumor reactions under spatiotemporally controllable conditions (Table 3). These applications together show the potential of sonogenetics in reshaping precision medicine.
Summary of therapeutic applications of sonogenetics in diverse disease models.
| Disease Models | Targets | Transducers | Mechanism | Therapeutic efficacy | Ref. |
|---|---|---|---|---|---|
| Parkinson's disease | STN | MscL-G22S | MscL-G22S activation excites STN neurons, modulating the specific brain circuit. | Improvement in balance and motor endurance. | [134] |
| Dopaminergic neurons of the SN | mPrestin (N7T, N308S) | Neuronal activation promotes expression of neurotrophic factors such as BDNF and NGF, thereby slowing the loss of TH+ neurons and restoring dopamine synthesis capacity. | After an 8-week course of treatment, dopaminergic neurodegeneration was reduced tenfold, and motor symptoms were alleviated fourfold. | [132] | |
| Epilepsy | GABAergic neurons | mPrestin (N7T, N308S) | Expressed in GABAergic neurons, mPrestin can be activated by 0.5 MHz transcranial ultrasound, causing these neurons to release GABA, which inhibits the abnormal synchronous excitation of nerve cells. | Upon ultrasound stimulation, epileptiform abnormal activity is directly reduced by 50%, and this inhibitory effect persists for 60 minutes. | [139] |
| Reward-related behavior | VTA | MscL-G22S | Activation of MscL in VTA neurons leads to dopamine release in the NAc (mesolimbic pathway activation). | Induction of appetitive conditioned reflex behavior. | [134] |
| Depressive-like behavior | DRN neurons | Endogenous sonosensitive mediators | Ultrasound mechanical forces amplified by PGVs can activate these channels, leading to rapid release of 5-HT. | Significant reduction in immobility time and prolongation of struggle time (TST/FST). | [10] |
| Visual restoration | V1 cortex | MscL-G22S | Activation of MscL triggers the generation of electrical signals in visual pathway neurons. These signals are decoded and processed by higher-order visual areas to form visual perception. | For mice expressing MscL, ultrasonic stimulation shows no statistically significant difference in success rates from the light-stimulated control group in associative learning tasks. | [6] |
| Tumor Therapy | Pancreatic cancer cells | Endogenous Piezo1 | Piezo1 is activated by external mechanical stimuli from US and MBs, mediating Ca²⁺ influx, inducing mitochondrial dysfunction, and ultimately leading to pancreatic cancer cell apoptosis. | Significantly delay tumor growth with US+MBs therapy. | [146] |
| Tumor cells | MscL I92L | Delivery of MscL I92L to tumor cells via cationic nanoliposomes, followed by ultrasonic activation of the channel, induces excessive Ca²⁺ influx and precisely triggers tumor cell apoptosis. | In mouse tumor models, the system successfully inhibits tumor growth and significantly improves survival rates. | [87] | |
| T cells (Jurkat T-cell lines/PBMC) | Endogenous Piezo1 | Piezo1 is activated by ultrasound + microbubbles, generating Ca²⁺ influx. Calcineurin, activated by Ca²⁺ influx, induces NFAT response elements, ultimately driving CAR gene expression. | Remote control of T-cell activation (CD69 upregulation) and CAR expression; enhanced cytotoxicity of ultrasound-induced PBMCs against target Nalm6 tumor cells. | [42] | |
| NSCLC A549 cells | MscL-I92G/I96G | Ultrasound activation of MscL induces calcium influx and cytoplasmic vacuolization, and decreases cell viability. | Reduced tumor volume and tumor weight. | [28] |
STN: subthalamic nucleus; TH: tyrosine hydroxylase; GABA: gamma-aminobutyric acid; VTA: ventral tegmental area; NAc: nucleus accumbens; DRN: dorsal raphe nucleus; PGVs: PEGylated gas vesicles; 5-HT: 5-hydroxytryptamine; TST: tail suspension test; FST: forced swim test; V1: primary visual cortex; US: ultrasound; MB: microbubble; CAR: chimeric antigen receptor; NFAT: nuclear factor of activated T-cells; PBMC: peripheral blood mononuclear cell; NSCLC: non-small cell lung cancer.
6.1 Neuromodulation
Neuromodulation aims to precisely regulate neural activity, which is crucial for the study of brain function and the treatment of functional disorders. Currently, the main methods of neural regulation include: deep brain stimulation (DBS), optogenetic interventions, transcranial magnetic stimulation (TMS), and transcranial direct current stimulation (tDCS). However, these techniques often have problems such as limited spatial resolution and penetration depth, or immune responses caused by invasiveness, which further restricts the application of some technologies [131]. Therefore, there is an urgent need for a noninvasive neuromodulation technology with high spatiotemporal accuracy and deep tissue penetration ability. Sonogenetics overcomes these limitations by using FUS to achieve deep tissue penetration and the use of mechanosensitive transducers that exhibit responsiveness to FUS [27].
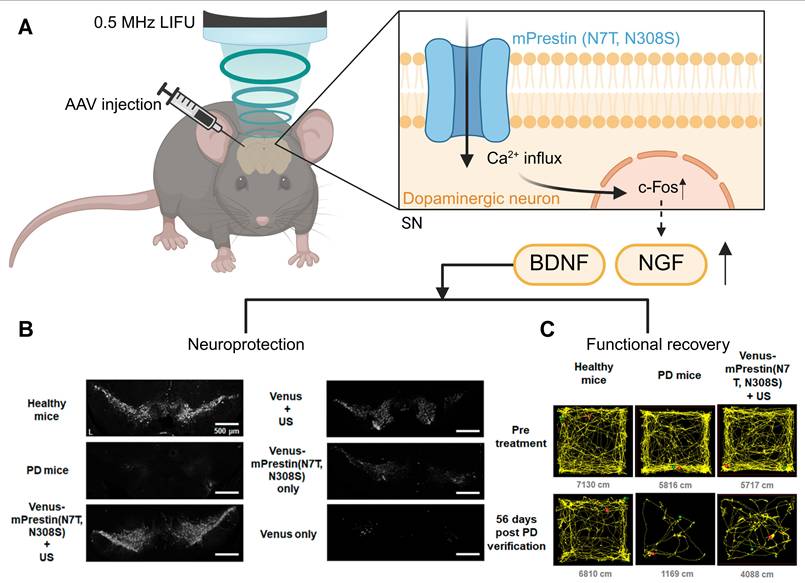
Sonogenetics can stimulate the deep brain without permanent implantation. It has a unique advantage in the treatment of neurodegenerative diseases. Parkinson's disease (PD) has motor and non-motor symptoms due to the degeneration of dopaminergic neurons [133]. Current treatment mainly focuses on symptom management. However, preclinical studies using sonogenetics have shown that this technology is expected to slow down the progression of the disease. In the Parkinson's disease mouse model, MscL-G22S was expressed in the subthalamic nuclei. Ultrasound-stimulated MscL-PD mice improved their balance and motor endurance through the modulation of specific brain circuits [134]. The dopaminergic neurons of the substantia nigra in PD mice expressed mPrestin (N7T, N308S) (Figure 6A). After ultrasound stimulation, the loss of TH-positive neurons in the substantia nigra (SN) significantly slowed down. In addition, repeated neuronal stimulation upregulated the expression of brain-derived neurotrophic factor (BDNF) and nerve growth factor (NGF), promoting neuroprotection and synaptic plasticity. Consequently, a significant enhancement of the motor function of PD mice was observed (Figure 6B, C) [132].
Schematic illustration of sonogenetic-based neuromodulation for the amelioration of Parkinson's disease. (A) The process of sonogenetics treats for Parkinson's disease. (B, C) This sonogenetic treatment results in neuroprotection, evidenced by preserved tyrosine hydroxylase-positive (TH⁺) neurons, and functional recovery, demonstrated by improved locomotor activity in open field tests. Adapted with permission from [132], Copyright 2021 American Chemical Society. Recreated with BioRender.com. AAV: adeno-associated virus; BDNF: brain-derived neurotrophic factor; c-Fos: cellular FBJ murine osteosarcoma viral oncogene homolog; LIFU: low-intensity focused ultrasound; NGF: nerve growth factor; PD: Parkinson’s disease; SN: substantia nigra; TH: tyrosine hydroxylase; US: ultrasound.
Epilepsy is a chronic brain dysfunction caused by abnormal synchronous excitation of neurons in the brain, manifested as convulsions and disturbances of consciousness. The present research generally has problems such as high invasiveness, insufficient targeting accuracy, and off-target effects [135-137]. Studies demonstrated that GABAergic neurons in the amygdala in the deep brain can inhibit neuronal excitation by specifically releasing gamma-aminobutyric acid (GABA), which is crucial for regulating epileptic seizures [138]. A study employing sonogenetics to treat epilepsy precisely harnessed this mechanism by expressing mPrestin (N7T, N308S) in GABAergic neurons. These neurons were activated via 0.5 MHz transcranial ultrasound, causing the release of GABA. GABA inhibited the abnormal synchronous excitation of nerve cells. In a rat epilepsy model induced by pentylenetetrazole, ultrasound stimulation directly reduced epileptiform abnormal activity by 50%, with the inhibitory effect persisting for 60 minutes. This study effectively suppresses epileptic seizures by ultrasonic remote control of GABA release [139]. Furthermore, sonogenetics plays a crucial role in modulating the reward circuitry of the ventral tegmental area (VTA). Under ultrasound stimulation, VTA neurons expressing MscL-G22S induced a rapid synchronous increase in DA signaling in the nucleus accumbens (NAc), thus regulating appetitive conditioning. In addition, the dorsal striatum (dSTR) expressing MscL-G22S can respond to ultrasound and increase locomotion in freely moving mice [134]. These studies demonstrate that sonogenetics has unique potential compared with traditional cerebral stimulation techniques.
In the research of endogenous sonosensitive mediators, PGVs+US stimulation in the dorsal raphe nucleus (DRN) was demonstrated to induce rapid release of 5-HT with exceptional temporal precision, effectively alleviating depression in mouse models [10]. While sonogenetic neuromodulation relying on engineered and exogenously expressed channels is undoubtedly important, the strategic harnessing of endogenous sonosensitive mediators within the cranium, such as Piezo1 in neurons [140] and TRPA1 in astrocytes [69], can eliminate dependence on gene delivery.
6.2 Vision restoration
Visual impairment severely compromises quality of life. Beyond conventional aids, gene therapies [141] and optogenetics have been demonstrated at the clinical level [142]. However, optogenetics is fundamentally limited by significant absorption and light scattering in brain tissue [143]. Sonogenetics takes the non-contact penetrative advantage of ultrasound to enable deep and long-range activation across the cortex. This approach is less invasive than current brain-machine interface (BMI) methods employed for visual restoration.
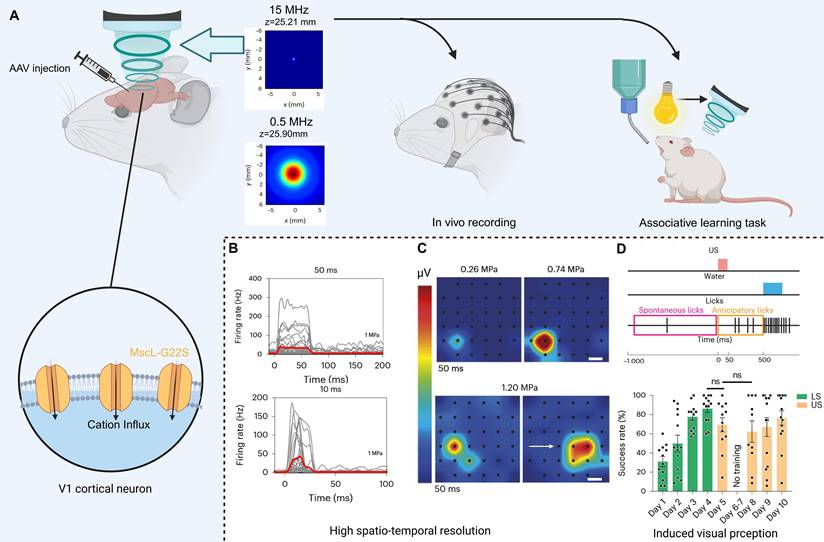
High spatiotemporal resolution is crucial for visual recovery to match visual frame rate and generate high-quality visual images. Toward this goal, Cadoni et al. demonstrated in both cellular and animal studies that sonogenetics provides the resolution necessary for vision restoration (Figure 7B, C). They chose the AAV to deliver MscL-G22S (Figure 7A). 15 MHz ultrasound stimulation evokes responses in the V1 cortex with the neural response latency under 10 ms. Moreover, the responses of the V1 cortex follow a repetition rate of up to 13 Hz that aligns with the transmission frequency required for visual recovery [144]. The spatial resolution of the V1 cortex is approximately 400 µm, compatible with the parameters required for visual restoration. Behavioral experiments further confirmed that 15 MHz ultrasonic stimuli can be perceived as visual by mice expressing MscL (Figure 7D). However, generating useful vision will necessitate sophisticated spatiotemporal patterning of ultrasound stimuli to construct coherent images, presenting a significant neural coding obstacle [6]. Whether the mechanical forces generated during treatment could damage the V1 cortex, and how ultrasound can reach the V1 cortex in human applications, are also issues that require attention.
High spatiotemporal resolution sonogenetic therapy for visual restoration using the MscL-G22S channel. (A) The treatment and experimental procedures for vision restoration induced by sonogenetics and comparison of acoustic beam profiles. (B) SDF shows millisecond-scale temporal precision in response to 10–50 ms pulses. (C) Pseudocolor activation maps reveal sub-millimeter spatial confinement that shifts precisely with transducer displacement. (D) In an associative learning task, mice expressing MscL-G22S in V1 perceive ultrasound stimulation as a visual cue. Adapted with permission from [6], copyright 2023 Springer Nature. Recreated with BioRender.com. AAV: adeno-associated virus; SDF: spike density function; V1: primary visual cortex.
6.3 Tumor therapy
Conventional cancer therapies like chemotherapy and radiotherapy lack sufficient precision, leading to systemic side effects [145]. Sonogenetics offers a novel strategy by integrating the deep penetration of focused ultrasound with genetically engineered mechanosensitivity. This approach triggers tumor-specific apoptosis while sparing healthy surrounding tissue.
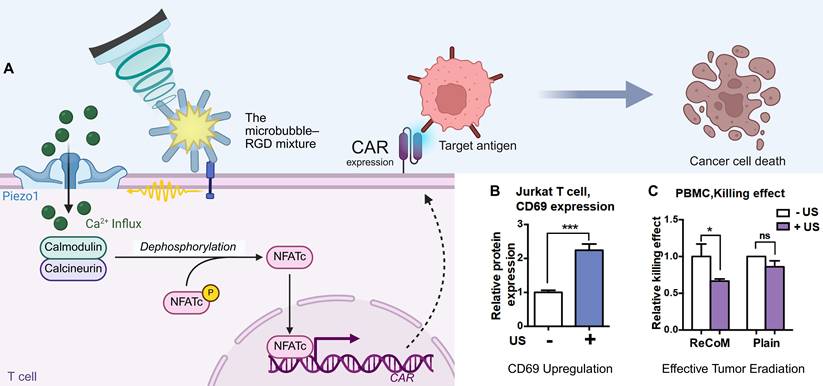
Current sonogenetic approaches in oncology can be categorized into three primary strategies. First, sonogenetics can directly suppress tumor growth. Piezo1 is highly expressed in the cell membrane of pancreatic cancer cell lines, particularly BxPC3. The activation of Piezo1, induced by external mechanical stimuli of ultrasound and MBs, mediates Ca²⁺ influx and mitochondrial dysfunction, ultimately leading to apoptosis in pancreatic cancer cells. In tumor xenograft models, US+MBs therapy significantly delayed tumor growth [146]. Alternatively, ultrasound has been shown to suppress the growth of tumors established from A549 cells that exogenously express MscL-I92G/I96G [28]. Second, sonogenetics can remotely control therapeutic immune cells. Ultrasound-activated Piezo1 in engineered T cells drives a calcium-dependent nuclear factor of activated T cells (NFAT) signaling pathway, which in turn induces the expression of chimeric antigen receptors (CARs) (Figure 8A). This enhances anti-tumor cytotoxicity in a spatiotemporally controlled manner (Figure 8B, C) [42]. Third, a novel "Logic AND-Gated Sonogene Nanosystem" was devised to precisely regulate tumor cell apoptosis. In this framework, LIFU (0.25 MPa) activates MscL I92L expressed in tumor cells to induce Ca2+ influx. When intracellular Ca2+ was excessive, it triggered the Ca2+ apoptosis pathway of cells. Experiments involving multiple tumor cell lines (HeLa, B16, 4T1) as well as in vivo treatment of the B16 tumor models, have demonstrated favorable tumor-suppressing effects [87]. The application of sonogenetics in the domain of tumor therapy has yielded a novel concept for precise, noninvasive eradication of tumor cells.
Mechanogenetics for remote control of cancer immunotherapy. (A) The schematic mechanism illustrates how sonogenetics induces the transcriptional expression of CAR. (B) The bar graphs show that ultrasound stimulation (+US) significantly upregulates the activation marker CD69 in Jurkat cells upon engagement with CD19⁺ tumor cells. (C) Luciferase-based killing assay shows that the ReCoM-engineered PBMCs demonstrate significant tumor killing efficacy upon ultrasound stimulation. Adapted with permission from [42], copyright 2018 National Academy of Sciences. Recreated with BioRender.com. CAR: chimeric antigen receptor; NFATc: nuclear factor of activated T-cells, cytoplasmic; PBMC: peripheral blood mononuclear cell; RGD: arginine-glycine-aspartic acid (peptide motif); US: ultrasound.
Similar to the synergy observed in the therapeutic application of the combination of sonogenetics and CAR-T, sonogenetics has the capacity to be integrated with conventional therapeutic modalities to confer spatiotemporal controllability. For example, combining sonogenetics with engineered bacteria allows for the localized, programmed release of drugs. Combining sonogenetics with systemic immune checkpoint inhibitors (e.g., anti-PD-1) is expected to overcome the immunosuppressive effect of the tumor microenvironment, enhancing the synergistic anti-tumor response. Drawing on the successful application of cell therapy in optogenetics [147], we propose to combine sonogenetics with stem cell technology: guiding embryonic stem cells (ESCs) or induced pluripotent stem cells (iPSCs) to differentiate into target cell types in vitro and undergo genetic engineering to stably express mechanosensitive proteins. Subsequently, these sonosensitive cells are transplanted into specific sites for targeted therapeutic intervention.
7. Conclusion and Future Outlook
Sonogenetics capitalizes on the comprehensive advantages of noninvasiveness, high spatiotemporal controllability, and deep tissue penetration. Compared to optogenetics, magnetogenetics, and chemogenetics (Table 4), it establishes a new paradigm for cellular regulation in precision medicine. This enables noninvasive treatment of deep-tissue diseases and mechanistic investigation, highlighting its irreplaceable value in precision medicine research [97,148]. In particular, for neuromodulation and the treatment of deep tissues, the penetrating power of ultrasound is unrivaled by technologies such as optogenetics. For instance, in the management of refractory epilepsy, sonogenetics can modulate deep-brain circuits without the requirement for invasive fiber-optic implants [139], thereby avoiding risks of neuroinflammation and glial scarring. For deep visceral tumors such as pancreatic cancer, the superior penetration of ultrasound overcomes the severe light-scattering limitations of optogenetics [70]. With the continuous development of portable ultrasound equipment, sonogenetics provides new methods for the long-term management of chronic diseases such as Parkinson's disease.
A comparative summary of sonogenetics, optogenetics, magnetogenetics, and chemogenetics.
| Technology | Toolkit | Mechanisms | Advantages | Limitations | Applications | Ref. |
|---|---|---|---|---|---|---|
| Sonogenetics | MS ion channels; Gene delivery vectors; Ultrasound | US-mediated mechanical effects (activation of mechanosensitive ion channels or macromolecular systems) or thermal effects (activation of thermoproteins or thermoresponsive promoters via elevated temperatures) enable remote regulation of cellular activity. | ·High spatial and temporal resolution; ·Noninvasive regulation | ·Off-target effect; ·Ultrasound parameter settings | ·Parkinson's disease ·Epilepsy; ·Neural circuits regulating; ·Visual restoration; ·Tumor therapy | [3,149] |
| Optogenetics | Photoresponsive proteins; Gene delivery vectors; Light delivery devices | Light delivery devices provide light of matching wavelength to activate photosensitive proteins. Upon receiving light signals, these photoreceptive proteins undergo conformational changes, triggering ion currents or protein interactions. | ·High spatial and temporal resolution; ·Strong cell type specificity | ·Low penetration depth; ·Invasive; ·Immune response and thermal damage from implanted devices | ·Retinitis pigmentosa; ·Deafness; ·Epilepsy; ·Parkinson's disease; ·Motor recovery after injury; ·Chronic pain; ·Cardiac arrhythmias; ·Laryngeal paralysis; ·Pancreatic islet-like organoid | [150,151] |
| Magnetogenetics | MNPs; Magnetic field | This technology exploits MNPs to induce mechanical or thermal stimuli within cells, thereby activating mechanosensitive and thermosensitive proteins. | ·Noninvasive regulation ·Multimodal | ·Precision of regulation; ·Clinical safety; ·Equipment condition; ·Unclear mechanism | ·Cancer therapy; ·Stem cell differentiation and tissue formation; ·Magnetically sensitive insulin-producing cells | [152,153] |
| Chemogenetics | DREADDs; CNO; AAV vectors | Through genetic engineering, receptors are designed to respond specifically to biologically inert ligands, thereby enabling remote regulation of cellular activity | ·High specificity; ·Duration of action | ·Low time accuracy; ·Side effects | ·Obesity; ·Sleep disorders; ·Epilepsy | [154-156] |
MS: mechanosensitive; US: ultrasound; AAV: adeno-associated virus; DREADDs: designer receptors exclusively activated by designer drugs; MNPs: magnetic nanoparticles; CNO: clozapine-N-oxide.
In order to facilitate the clinical translation of sonogenetics, there are several critical issues that must be addressed. The lack of a unified standard for frequency, intensity, and duty cycle hinders cross-study comparisons of distinct mechanosensitive transducers. Improper parameter settings can either reduce the regulatory efficiency or trigger adverse thermal effects and inertial cavitation, leading to biological tissue damage [157]. Second, because few studies have investigated the distribution and responsiveness of endogenous MS ion channels across different tissues, activating exogenously expressed target MS ion channels may inadvertently affect non-target cells [158,159]. Such off-target activation impairs the reliability of sonogenetics as a precision regulatory tool. Microbubbles designed to target specific cells at the molecular level [10,43] are expected to address this issue. Given the off-target effects of endogenous channels, we recommend prioritizing the selection of exogenous channels that offer high specificity for high-precision clinical applications. Furthermore, achieving bidirectional and precise control over cellular activity remains a significant challenge. To date, among the studies of MS channels, only the K2P channel family (such as TRAAK, TREK-1, TREK-2) exhibits K⁺ efflux upon activation, producing hyperpolarizing effect. The identification and introduction of additional inhibitory channels, such as K⁺ or Cl⁻ channels, are critical for expanding the sonogenetic toolkit. Additionally, as sonogenetics relies heavily on effective gene delivery, the inherent limitations of this delivery process pose a critical challenge. In practical clinical settings, we must consider potential pharmacological interference. Other drugs that the patient is taking, such as calcium channel blockers or neuromodulators, may inadvertently change the resting membrane potential or gate threshold of the target MS channels. Therefore, when selecting ultrasound parameters, the specific situation of the patient should be fully considered.
Sonogenetics is a multidisciplinary field. To promote the clinical translation of sonogenetics, interdisciplinary cooperation is important. Therefore, we present the following strategic roadmap organized by discipline. In molecular biology, we modify channels to achieve heightened ultrasonic sensitivity and optimize gene delivery protocols. Researchers should continue to explore novel MS ion channels and their mutants to improve mechanical sensitivity and achieve faster response speed. Furthermore, the biophysical properties of known MS ion channels, such as activation thresholds and biosafety, must be comprehensively characterized. Introduce certain well-researched MS ion channels, such as MscL-G22S, into non-human primates to further verify their safety and effectiveness under conditions closer to human environments. In engineering, we continuously refine the focusing precision of ultrasonic devices and develop lightweight and portable equipment for long-term management of chronic diseases. CNS represents a key application area of sonogenetics. Noninvasive ultrasound devices capable of penetrating the skull and delivering precise focusing in humans require further development in order to fully realize the benefits of sonogenetics in terms of precision and noninvasiveness. In clinical medicine, standardized clinical workflows must be established in accordance with FDA guidelines (MI, ISPTA). The determination of ultrasound parameters, imaging monitoring of MS ion channel expression and localization, and the establishment of standardized metrics should be integrated to form a closed-loop regulatory and efficacy assessment system. Moreover, doctors can design safe sonogenetic approaches tailored to disease characteristics. Finally, we can draw valuable experience from analogous technologies to advance sonogenetics. Future research should explore multimodal synergies to combine the strengths of different modalities. For instance, sono-optogenetics increases the penetration depth [160] and autophagy is regulated through combined optogenetics and chemogenetics [161]. The construction of such a multimodal system may provide some means to overcome current technological limitations.
In essence, sonogenetics resolves the long-standing conflict between "precision" and "noninvasiveness" in precision medicine. It is expected to change the treatment pattern of incurable diseases such as neurodegenerative disorders, tumors, and visual impairments. Through systematic optimization tools, the establishment of standardized workflows, and strict clinical verification, sonogenetics will become a cornerstone of noninvasive precision medicine, bringing new hope to patients with unmet clinical needs currently.
Abbreviations
5-HT: 5-Hydroxytryptamine; AAV: adeno-associated virus; AdV: Adenovirus; ARF: acoustic radiation force; ARG: acoustic reporter gene; ASIC: acid-sensing ion channel; ATC: acoustic tweezing cytometry; BBB: blood-brain barrier; BDNF: brain-derived neurotrophic factor; BMI: brain-machine interface; BURST: burst ultrasound reconstructed with signal templates; CAR: chimeric antigen receptor; CNO: clozapine-N-oxide; CNS: central nervous system; DEG/ENaC: degenerin/epithelial Na+ channel; DREADDs: designer receptors exclusively activated by designer drugs; DRN: dorsal raphe nucleus; dSTR: dorsal striatum; ESC: embryonic stem cell; FUS: Focused Ultrasound; GABA: gamma-aminobutyric acid; GFP: green fluorescent protein; GV: Gas Vesicle; HEK 293T: human embryonic kidney 293T; hSyn: human synapsin; iPSC: Induced Pluripotent Stem Cell; ISATA: spatial average temporal average intensity; ISPTA: spatial peak temporal average intensity; K2P: two-pore domain potassium channel; LIFU: low-intensity focused ultrasound; LNP: lipid nanoparticle; LV: lentivirus; MB: microbubble; MNP: Magnetic Nanoparticle; MRI: magnetic resonance imaging; MRgFUS: magnetic resonance-guided focused ultrasound surgery; MS (channels): Mechanosensitive (channels); MscL: large-conductance mechanosensitive channel; NAc: nucleus accumbens; NFAT: nuclear factor of activated T-cells; NGF: nerve growth factor; NSCLC: non-small cell lung cancer; PBMC: peripheral blood mononuclear cell; PD: Parkinson's Disease; PEG: Polyethylene Glycol; PGVs: PEGylated Gas Vesicles; PTMB: Piezo1-targeted microbubble; SAW: Surface acoustic wave; SN: substantia nigra; STN: subthalamic nucleus; tDCS: transcranial direct current stimulation; TMS: transcranial magnetic stimulation; UMGAAV: ultrasound and microbubble guided adenoassociated viral vector; US: Ultrasound; V1: primary visual cortex; VTA: ventral tegmental area; WT: wild-type; YAP: yes-associated protein; YFP: yellow fluorescent protein.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (No. 8257071094).
We thank BioRender (biorender.com) for creation of Figures 1, 3–8. We also acknowledge the assistance of Gemini 3.1 pro for language polishing and grammatical refinement, but not for any content generation.
Author contributions
Xinrui Yu: Data acquisition, Writing – original draft, Resources. Min Pu: Conceptualization, Writing – review and editing. Ruoli Wang: Writing – review and editing, Visualization. Haitao Ran: Supervision, Project administration, Funding acquisition, Writing – review and editing.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Tang J, Feng M, Wang D, Zhang L, Yang K. Recent advancement of sonogenetics: a promising noninvasive cellular manipulation by ultrasound. Genes Dis. 2024;11:101112
2. Wu Y, Liu Y, Huang Z, Wang X, Jin Z, Li J. et al. Control of the activity of CAR-T cells within tumours via focused ultrasound. Nat Biomed Eng. 2021;5:1336-47
3. Wu P, Liu Z, Tao W, Lai Y, Yang G, Yuan L. The principles and promising future of sonogenetics for precision medicine. Theranostics. 2024;14:4806-21
4. Liu P, Foiret J, Situ Y, Zhang N, Kare AJ, Wu B. et al. Sonogenetic control of multiplexed genome regulation and base editing. Nat Commun. 2023;14:6575
5. Huang D, Wang J, Song C, Zhao Y. Ultrasound-responsive matters for biomedical applications. Innovation (Camb). 2023;4:100421
6. Cadoni S, Demené C, Alcala I, Provansal M, Nguyen D, Nelidova D. et al. Ectopic expression of a mechanosensitive channel confers spatiotemporal resolution to ultrasound stimulations of neurons for visual restoration. Nat Nanotechnol. 2023;18:667-76
7. Ashar H, Ranjan A. Immunomodulation and targeted drug delivery with high intensity focused ultrasound (HIFU): principles and mechanisms. Pharmacol Ther. 2023;244:108393
8. Rao V, Coetzee J, Parvaz L, Penalba A, Adamson M, Yoon BC. Evaluation of temperature elevation associated with low-intensity focused ultrasound (LIFU) stimulation with or without a deep brain electrode. Brain Stimul. 2025;18:1876-8
9. Ibsen S, Tong A, Schutt C, Esener S, Chalasani SH. Sonogenetics is a non-invasive approach to activating neurons in caenorhabditis elegans. Nat Commun. 2015;6:8264
10. Hou X, Jing J, Jiang Y, Huang X, Xian Q, Lei T. et al. Nanobubble-actuated ultrasound neuromodulation for selectively shaping behavior in mice. Nat Commun. 2024;15:2253
11. Legon W, Strohman A. Low-intensity focused ultrasound for human neuromodulation. Nat Rev Methods Primer. 2024;4:91
12. Qiu Z, Kala S, Guo J, Xian Q, Zhu J, Zhu T. et al. Targeted neurostimulation in mouse brains with non-invasive ultrasound. Cell Rep. 2020;32:108033
13. Chen S, Nazeri A, Baek H, Ye D, Yang Y, Yuan J. et al. A review of bioeffects induced by focused ultrasound combined with microbubbles on the neurovascular unit. J Cereb Blood Flow Metab. 2022;42:3-26
14. Du J, Liao M, Zhang D, Li X. Advanced strategies for ultrasound control and applications in sonogenetics and gas vesicle-based technologies: a review. Int J Nanomedicine. 2025;20:6533-49
15. Schoen S, Kilinc MS, Lee H, Guo Y, Degertekin FL, Woodworth GF. et al. Towards controlled drug delivery in brain tumors with microbubble-enhanced focused ultrasound. Adv Drug Deliv Rev. 2022;180:114043
16. Paranjape AN, D’Aiuto L, Zheng W, Ramasamy T, Chen X, Villanueva FS. Abstract 13414: studying sonoporation-induced drug payload penetration beyond the blood-brain barrier using multi-cellular human brain spheroids. Circulation. 2021;144:13414
17. Wang D, Xing C, Liang Y, Wang C, Zhao P, Liang X. et al. Ultrasound imaging of tumor vascular CD93 with MMRN2 modified microbubbles for immune microenvironment prediction. Adv Mater. 2024;36:e2310421
18. Pirovano G, Roberts S, Kossatz S, Reiner T. Optical imaging modalities: principles and applications in preclinical research and clinical settings. J Nucl Med. 2020;61:1419-27
19. Poole K. The diverse physiological functions of mechanically activated ion channels in mammals. Annu Rev Physiol. 2022;84:307-29
20. Kefauver JM, Ward AB, Patapoutian A. Discoveries in structure and physiology of mechanically activated ion channels. Nature. 2020;587:567-76
21. Wang J, Blount P, Hou T, Sokabe M. Editorial: In silico gating mechanism studies and modulator discovery for MscL. Front Chem. 2024;12:1376617
22. Walton TA, Idigo CA, Herrera N, Rees DC. MscL: channeling membrane tension. Pflugers Arch. 2015;467:15-25
23. Bavi N, Cortes DM, Cox CD, Rohde PR, Liu W, Deitmer JW. et al. The role of MscL amphipathic N terminus indicates a blueprint for bilayer-mediated gating of mechanosensitive channels. Nat Commun. 2016;7:11984
24. Chang G, Spencer RH, Lee AT, Barclay MT, Rees DC. Structure of the MscL homolog from mycobacterium tuberculosis: a gated mechanosensitive ion channel. Science. 1998;282:2220-6
25. Heureaux J, Chen D, Murray VL, Deng CX, Liu AP. Activation of a bacterial mechanosensitive channel in mammalian cells by cytoskeletal stress. Cell Mol Bioeng. 2014;7:307-19
26. Li X, Zheng C, Tian Y, Ming D. Channel-specific differential effects of bacterial mechanosensitive channels for ultrasound neuromodulation in precision sonogenetics. Theranostics. 2026;16:2447-65
27. Ye J, Tang S, Meng L, Li X, Wen X, Chen S. et al. Ultrasonic control of neural activity through activation of the mechanosensitive channel MscL. Nano Lett. 2018;18:4148-55
28. Wen X, Wang Y, Zhu Z, Guo S, Qian J, Zhu J. et al. Mechanosensitive channel MscL induces non-apoptotic cell death and its suppression of tumor growth by ultrasound. Front Chem. 2023;11:1130563
29. Kubanek J, Shi J, Marsh J, Chen D, Deng C, Cui J. Ultrasound modulates ion channel currents. Sci Rep. 2016;6:24170
30. Zhang M, Ma Y, Ye X, Zhang N, Pan L, Wang B. TRP (transient receptor potential) ion channel family: structures, biological functions and therapeutic interventions for diseases. Signal Transduct Target Ther. 2023;8:261
31. Samanta A, Hughes TET, Moiseenkova-Bell VY. Transient receptor potential (TRP) channels. Subcell Biochem. 2018;87:141-65
32. Ramsey IS, Delling M, Clapham DE. An introduction to TRP channels. Annu Rev Physiol. 2006;68:619-47
33. Kang L, Gao J, Schafer WR, Xie Z, Xu XZS. C. elegans TRP family protein TRP-4 is a pore-forming subunit of a native mechanotransduction channel. Neuron. 2010;67:381-91
34. Magaram U, Weiss C, Vasan A, Reddy KC, Friend J, Chalasani SH. Two pathways are required for ultrasound-evoked behavioral changes in caenorhabditis elegans. PLoS One. 2022;17:e0267698
35. Yoo S, Mittelstein DR, Hurt RC, Lacroix J, Shapiro MG. Focused ultrasound excites cortical neurons via mechanosensitive calcium accumulation and ion channel amplification. Nat Commun. 2022;13:493
36. Duque M, Lee-Kubli CA, Tufail Y, Magaram U, Patel J, Chakraborty A. et al. Sonogenetic control of mammalian cells using exogenous Transient Receptor Potential A1 channels. Nat Commun. 2022;13:1130
37. Matsushita Y, Yoshida K, Yoshiya M, Shimizu T, Tsukamoto S, Kudo N. et al. TRPC6 is a mechanosensitive channel essential for ultrasound neuromodulation in the mammalian brain. Proc Natl Acad Sci U S A. 2024;121:e2404877121
38. Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS. et al. Piezo proteins are pore-forming subunits of mechanically activated channels. Nature. 2012;483:176-81
39. Jiang Y, Yang X, Jiang J, Xiao B. Structural designs and mechanogating mechanisms of the mechanosensitive piezo channels. Trends Biochem Sci. 2021;46:472-88
40. Zhao Q, Wu K, Geng J, Chi S, Wang Y, Zhi P. et al. Ion Permeation and Mechanotransduction Mechanisms of Mechanosensitive Piezo Channels. Neuron. 2016;89:1248-63
41. Xiao B. Mechanisms of mechanotransduction and physiological roles of PIEZO channels. Nat Rev Mol Cell Biol. 2024;25:886-903
42. Pan Y, Yoon S, Sun J, Huang Z, Lee C, Allen M. et al. Mechanogenetics for the remote and noninvasive control of cancer immunotherapy. Proc Natl Acad Sci U S A. 2018;115:992-7
43. Shen X, Song Z, Xu E, Zhou J, Yan F. Sensitization of nerve cells to ultrasound stimulation through Piezo1-targeted microbubbles. Ultrason Sonochem. 2021;73:105494
44. Jiang Z, Chen Z, Xu Y, Li H, Li Y, Peng L. et al. Low-frequency ultrasound sensitive Piezo1 channels regulate keloid-related characteristics of fibroblasts. Adv Sci Weinh Baden-Wurtt Ger. 2024;11:e2305489
45. Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ. et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science. 2010;330:55-60
46. Lolli G, Pasqualetto E, Costanzi E, Bonetto G, Battistutta R. The STAS domain of mammalian SLC26A5 prestin harbours an anion-binding site. Biochem J. 2016;473:365-70
47. Liu XZ. Prestin, a cochlear motor protein, is defective in non-syndromic hearing loss. Hum Mol Genet. 2003;12:1155-62
48. Schänzler M, Fahlke C. Anion transport by the cochlear motor protein prestin. J Physiol. 2011;590:259-72
49. Santos-Sacchi J, Bai J-P, Navaratnam D. Megahertz Sampling of Prestin (SLC26a5) Voltage-Sensor Charge Movements in Outer Hair Cell Membranes Reveals Ultrasonic Activity that May Support Electromotility and Cochlear Amplification. J Neurosci. 2023;43:2460-8
50. Huang Y-S, Fan C-H, Hsu N, Chiu N-H, Wu C-Y, Chang C-Y. et al. Sonogenetic modulation of cellular activities using an engineered auditory-sensing protein. Nano Lett. 2020;20:1089-100
51. Wu C-Y, Fan C-H, Chiu N-H, Ho Y-J, Lin Y-C, Yeh C-K. Targeted delivery of engineered auditory sensing protein for ultrasound neuromodulation in the brain. Theranostics. 2020;10:3546-61
52. Wang H-C, Phan T-N, Chen T-H, Lin Y, Hsu N, Fan C-H. et al. Enhancing prestin oligomerization for superior sonogenetics. Theranostics. 2026;16:3321-37
53. Schewe M, Nematian-Ardestani E, Sun H, Musinszki M, Cordeiro S, Bucci G. et al. A non-canonical voltage-sensing mechanism controls gating in K2P K(+) channels. Cell. 2016;164:937-49
54. Brohawn SG, Su Z, MacKinnon R. Mechanosensitivity is mediated directly by the lipid membrane in TRAAK and TREK1 K channels. Proc Natl Acad Sci U S A. 2014;111:3614-9
55. Kang D, Choe C, Kim D. Thermosensitivity of the two-pore domain K channels TREK-2 and TRAAK. J Physiol. 2005;564:103-16
56. Maingret F, Patel AJ, Lesage F, Lazdunski M, Honoré E. Mechano- or acid stimulation, two interactive modes of activation of the TREK-1 potassium channel. J Biol Chem. 1999;274:26691-6
57. Lolicato M, Arrigoni C, Mori T, Sekioka Y, Bryant C, Clark KA. et al. K2P2.1 (TREK-1)-activator complexes reveal a cryptic selectivity filter binding site. Nature. 2017;547:364-8
58. Brohawn SG. How ion channels sense mechanical force: insights from mechanosensitive K2P channels TRAAK, TREK1, and TREK2. Ann N Y Acad Sci. 2015;1352:20-32
59. Sorum B, Docter T, Panico V, Rietmeijer RA, Brohawn SG. Tension activation of mechanosensitive two-pore domain K+ channels TRAAK, TREK-1, and TREK-2. Nat Commun. 2024;15:3142
60. Douguet D, Honoré E. Mammalian Mechanoelectrical Transduction: Structure and Function of Force-Gated Ion Channels. Cell. 2019;179:340-54
61. Sorum B, Rietmeijer RA, Gopakumar K, Adesnik H, Brohawn SG. Ultrasound activates mechanosensitive TRAAK K+ channels through the lipid membrane. Proc Natl Acad Sci U S A. 2021;118:e2006980118
62. Herrera-Pérez S, Campos-Ríos A, Rueda-Ruzafa L, Lamas JA. Contribution of K2P potassium channels to cardiac physiology and pathophysiology. Int J Mol Sci. 2021;22:6635
63. Lim J, Tai H-H, Liao W-H, Chu Y-C, Hao C-M, Huang Y-C. et al. ASIC1a is required for neuronal activation via low-intensity ultrasound stimulation in mouse brain. Elife. 2021;10:e61660
64. Han DS, Chen CC. Aspirin, an ASIC3 inhibitor, blocks the analgesic effect of therapeutic ultrasound in chronic muscle pain. Ann Phys Rehabil Med. 2018;61:e420-1
65. Puts R, Rikeit P, Ruschke K, Knaus P, Schreivogel S, Raum K. Functional regulation of YAP mechanosensitive transcriptional coactivator by focused low-intensity pulsed ultrasound (FLIPUS) enhances proliferation of murine mesenchymal precursors. PLoS One. 2018;13:e0206041
66. Driscoll M, Chalfie M. The mec-4 gene is a member of a family of caenorhabditis elegans genes that can mutate to induce neuronal degeneration. Nature. 1991;349:588-93
67. Chelur DS, Ernstrom GG, Goodman MB, Yao CA, Chen L, Hagan RO. et al. The mechanosensory protein MEC-6 is a subunit of the C. elegans touch-cell degenerin channel. Nature. 2002;420:669-73
68. Zhou W, Wang X, Wang K, Farooq U, Kang L, Niu L. et al. Ultrasound activation of mechanosensory ion channels in caenorhabditis elegans. IEEE Trans Ultrason Ferroelectr Freq Control. 2022;69:473-9
69. Oh S-J, Lee JM, Kim H-B, Lee J, Han S, Bae JY. et al. Ultrasonic neuromodulation via astrocytic TRPA1. Curr Biol. 2019;29:3386-401.e8
70. Zhu M, Fang Y, Sun Y, Li S, Yu J, Xiong B. et al. Sonogenetics in the treatment of chronic diseases: a new method for cell regulation. Adv Sci (Weinh). 2024;11:e2407373
71. Naso MF, Tomkowicz B, Perry WL, Strohl WR. Adeno-associated virus (AAV) as a vector for gene therapy. BioDrugs. 2017;31:317-34
72. Russell S, Bennett J, Wellman JA, Chung DC, Yu Z-F, Tillman A. et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65-mediated inherited retinal dystrophy: a randomised, controlled, open-label, phase 3 trial. Lancet. 2017;390:849-60
73. Mehta N, Gilbert R, Chahal PS, Moreno MJ, Nassoury N, Coulombe N. et al. Preclinical development and characterization of novel adeno-associated viral vectors for the treatment of lipoprotein lipase deficiency. Hum Gene Ther. 2023;34:927-46
74. Chu D, Sullivan CC, Weitzman MD, Du L, Wolf PL, Jamieson SW. et al. Direct comparison of efficiency and stability of gene transfer into the mammalian heart using adeno-associated virus versus adenovirus vectors. J Thorac Cardiovasc Surg. 2003;126:671-9
75. Chandler RJ, LaFave MC, Varshney GK, Trivedi NS, Carrillo-Carrasco N, Senac JS. et al. Vector design influences hepatic genotoxicity after adeno-associated virus gene therapy. J Clin Invest. 2015;125:870-80
76. Kofoed RH, Dibia CL, Noseworthy K, Xhima K, Vacaresse N, Hynynen K. et al. Efficacy of gene delivery to the brain using AAV and ultrasound depends on serotypes and brain areas. J Control Release. 2022;351:667-80
77. Li C, Samulski RJ. Engineering adeno-associated virus vectors for gene therapy. Nat Rev Genet. 2020;21:255-72
78. Kay MA, Glorioso JC, Naldini L. Viral vectors for gene therapy: the art of turning infectious agents into vehicles of therapeutics. Nat Med. 2001;7:33-40
79. Wanisch K, Yáñez-Muñoz RJ. Integration-deficient lentiviral vectors: a slow coming of age. Mol Ther. 2009;17:1316-32
80. Jargalsaikhan B-E, Muto M, Ema M. The era of gene therapy: the advancement of lentiviral vectors and their pseudotyping. Viruses. 2025;17:1036
81. Hislop JN, Islam TA, Eleftheriadou I, Carpentier DCJ, Trabalza A, Parkinson M. et al. Rabies virus envelope glycoprotein targets lentiviral vectors to the axonal retrograde pathway in motor neurons. J Biol Chem. 2014;289:16148-63
82. Höfig I, Barth S, Salomon M, Jagusch V, Atkinson MJ, Anastasov N. et al. Systematic improvement of lentivirus transduction protocols by antibody fragments fused to VSV-G as envelope glycoprotein. Biomaterials. 2014;35:4204-12
83. Lee CS, Bishop ES, Zhang R, Yu X, Farina EM, Yan S. et al. Adenovirus-mediated gene delivery: potential applications for gene and cell-based therapies in the new era of personalized medicine. Genes Dis. 2017;4:43-63
84. Raper SE, Chirmule N, Lee FS, Wivel NA, Bagg A, Gao G. et al. Fatal systemic inflammatory response syndrome in a ornithine transcarbamylase deficient patient following adenoviral gene transfer. Mol Genet Metab. 2003;80:148-58
85. Gillmore JD, Gane E, Taubel J, Kao J, Fontana M, Maitland ML. et al. CRISPR-Cas9 In vivo gene editing for transthyretin amyloidosis. N Engl J Med. 2021;385:493-502
86. Kenjo E, Hozumi H, Makita Y, Iwabuchi KA, Fujimoto N, Matsumoto S. et al. Low immunogenicity of LNP allows repeated administrations of CRISPR-Cas9 mRNA into skeletal muscle in mice. Nat Commun. 2021;12:7101
87. Wang T, Wang H, Pang G, He T, Yu P, Cheng G. et al. A Logic AND-Gated Sonogene Nanosystem for Precisely Regulating the Apoptosis of Tumor Cells. ACS Appl Mater Interfaces. 2020;12:56692-700
88. Song K-H, Harvey BK, Borden MA. State-of-the-art of microbubble-assisted blood-brain barrier disruption. Theranostics. 2018;8:4393-408
89. Tung Y-S, Vlachos F, Feshitan JA, Borden MA, Konofagou EE. The mechanism of interaction between focused ultrasound and microbubbles in blood-brain barrier opening in mice. J Acoust Soc Am. 2011;130:3059-67
90. Chien C-Y, Xu L, Pacia CP, Yue Y, Chen H. Blood-brain barrier opening in a large animal model using closed-loop microbubble cavitation-based feedback control of focused ultrasound sonication. Sci Rep. 2022;12:16147
91. Cattaneo M, Guerriero G, Shakya G, Krattiger LA, Paganella LG, Narciso ML. et al. Cyclic jetting enables microbubble-mediated drug delivery. Nat Phys. 2025;21:590-8
92. Zhu Y, Arkin G, Zeng W, Huang Y, Su L, Guo F. et al. Ultrasound image-guided cancer gene therapy using iRGD dual-targeted magnetic cationic microbubbles. Biomed Pharmacother. 2024;172:116221
93. Azmin M, Harfield C, Ahmad Z, Edirisinghe M, Stride E. How do microbubbles and ultrasound interact? Basic physical, dynamic and engineering principles. Curr Pharm Des. 2012;18:2118-34
94. Challis RC, Kumar SR, Chan KY, Challis C, Beadle K, Jang MJ. et al. Systemic AAV vectors for widespread and targeted gene delivery in rodents. Nat Protoc. 2019;14:379-414
95. Yin L, He H, Zhang H, Shang Y, Fu C, Wu S. et al. Revolution of AAV in drug discovery: from delivery system to clinical application. J Med Virol. 2025;97:e70447
96. Chen J, Zhan L, Lv C, Teng J, Cao C. Advances and challenges in adeno-associated virus gene therapy applications of localized delivery strategies. Curr Med Sci. 2025;45:683-98
97. Geng G, Xu Y, Hu Z, Wang H, Chen X, Yuan W. et al. Viral and non-viral vectors in gene therapy: current state and clinical perspectives. EBioMedicine. 2025;118:105834
98. Gouvarchin Ghaleh HE, Bolandian M, Dorostkar R, Jafari A, Pour MF. Concise review on optimized methods in production and transduction of lentiviral vectors in order to facilitate immunotherapy and gene therapy. Biomed Pharmacother. 2020;128:110276
99. Richter M, Saydaminova K, Yumul R, Krishnan R, Liu J, Nagy EE. et al. In vivo transduction of primitive mobilized hematopoietic stem cells after intravenous injection of integrating adenovirus vectors. Blood. 2016;128:2206-2217
100. Boucher P, Cui X, Curiel DT. Adenoviral vectors for in vivo delivery of CRISPR-cas gene editors. J Control Release. 2020;327:788-800
101. Raguram A, Banskota S, Liu DR. Therapeutic in vivo delivery of gene editing agents. Cell. 2022;185:2806-27
102. Jeong M, Lee Y, Park J, Jung H, Lee H. Lipid nanoparticles (LNPs) for in vivo RNA delivery and their breakthrough technology for future applications. Adv Drug Deliv Rev. 2023;200:114990
103. Jallow MB, Huang K, Qiu M. Versatility of LNPs across different administration routes for targeted RNA delivery. J Mater Chem B. 2025;13:7637-52
104. Kim K, Lee J, Park M-H. Microbubble delivery platform for ultrasound-mediated therapy in brain cancers. Pharmaceutics. 2023;15:698
105. Xie A, Wu MD, Cigarroa G, Belcik JT, Ammi A, Moccetti F. et al. Influence of DNA-microbubble coupling on contrast ultrasound-mediated gene transfection in muscle and liver. J Am Soc Echocardiogr. 2016;29:812-8
106. Tie Z, Zhang S, Qu Y, Wang M, Liu R, Sun D. et al. Advances in microbubble-assisted ultrasound-guided gene therapy: mechanisms and applications. Sci China Mater. 2024;67:2749-66
107. Hou X, Qiu Z, Xian Q, Kala S, Jing J, Wong KF. et al. Precise ultrasound neuromodulation in a deep brain region using nano gas vesicles as actuators. Adv Sci. 2021;8:e2101934
108. Ennasr A, Isaac G, Strohman A, Legon W. Examination of the interaction of parameters for low-intensity focused ultrasound of the human motor cortex. Brain Stimul. 2024;17:1293-306
109. Sarica C, Nankoo J-F, Fomenko A, Grippe TC, Yamamoto K, Samuel N. et al. Human studies of transcranial ultrasound neuromodulation: a systematic review of effectiveness and safety. Brain Stimul. 2022;15:737-746
110. Shurche S, Dizaji MM, Movahedin M, Nilforoushzadeh MA, Taghiabadi E. Mechanical index-guided low intensity ultrasound therapy for hair regeneration: insights from finite element modeling and in vivo experiments. Phys Fluids. 2025;37:117114
111. Duck FA. Medical and non-medical protection standards for ultrasound and infrasound. Prog Biophys Mol Biol. 2007;93:176-91
112. Fry FJ, Barger JE. Acoustical properties of the human skull. J Acoust Soc Am. 1978;63:1576-90
113. Pichardo S, Sin VW, Hynynen K. Multi-frequency characterization of the speed of sound and attenuation coefficient for longitudinal transmission of freshly excised human skulls. Phys Med Biol. 2011;56:219-50
114. Clement GT, Hynynen K. A non-invasive method for focusing ultrasound through the human skull. Phys Med Biol. 2002;47:1219-36
115. Tang KWK, Jeong J, Hsieh J-C, Yao M, Ding H, Wang W. et al. Bioadhesive hydrogel-coupled and miniaturized ultrasound transducer system for long-term, wearable neuromodulation. Nat Commun. 2025;16:4940
116. Yoshimura K, Batiza A, Schroeder M, Blount P, Kung C. Hydrophilicity of a single residue within MscL correlates with increased channel mechanosensitivity. Biophys J. 1999;77:1960-72
117. Yoshimura K, Usukura J, Sokabe M. Gating-associated conformational changes in the mechanosensitive channel MscL. Proc Natl Acad Sci U S A. 2008;105:4033-8
118. Li Y, Wray R, Eaton C, Blount P. An open-pore structure of the mechanosensitive channel MscL derived by determining transmembrane domain interactions upon gating. FASEB J. 2009;23:2197-204
119. Catane R, Beck A, Inbar Y, Rabin T, Shabshin N, Hengst S. et al. MR-guided focused ultrasound surgery (MRgFUS) for the palliation of pain in patients with bone metastases-preliminary clinical experience. Ann Oncol. 2007;18:163-7
120. Meng Y, Reilly RM, Pezo RC, Trudeau M, Sahgal A, Singnurkar A. et al. MR-guided focused ultrasound enhances delivery of trastuzumab to Her2-positive brain metastases. Sci Transl Med. 2021;13:eabj4011
121. Blesa J, Pineda-Pardo JA, Inoue KI, Gasca-Salas C, Balzano T, Del Rey NL. et al. BBB opening with focused ultrasound in nonhuman primates and Parkinson's disease patients: targeted AAV vector delivery and PET imaging. Sci Adv. 2023;9:eadf4888
122. Donfrancesco AD, Adelizzi A, Giri A, Duchi R, Boito S, Barandalla M. et al. Transabdominal ultrasound guided AAV9-GFP delivery in fetal pigs: a translational and minimally invasive model for in utero fetal gene therapy. Gene Ther. 2025;32:487-96
123. Liu R, Qu S, Xu Y, Jo H, Dai Z. Spatial control of robust transgene expression in mouse artery endothelium under ultrasound guidance. Signal Transduct Target Ther. 2022;7:225
124. Concilio SC, Russell SJ, Peng K-W. A brief review of reporter gene imaging in oncolytic virotherapy and gene therapy. Mol Ther Oncol. 2021;21:98-109
125. Serganova I, Blasberg RG. Molecular imaging with reporter genes: has its promise been delivered? J Nucl Med. 2019;60:1665-81
126. Wu ZJ, Tang FR, Ma Z-W, Peng X-C, Xiang Y, Zhang Y. et al. Oncolytic viruses for tumor precision imaging and radiotherapy. Hum Gene Ther. 2018;29:204-22
127. Bourdeau RW, Lee-Gosselin A, Lakshmanan A, Farhadi A, Kumar SR, Nety SP. et al. Acoustic reporter genes for noninvasive imaging of microorganisms in mammalian hosts. Nature. 2018;553:86-90
128. Farhadi A, Ho GH, Sawyer DP, Bourdeau RW, Shapiro MG. Ultrasound imaging of gene expression in mammalian cells. Science. 2019;365:1469-75
129. Sawyer DP, Bar-Zion A, Farhadi A, Shivaei S, Ling B, Lee-Gosselin A. et al. Ultrasensitive ultrasound imaging of gene expression with signal unmixing. Nat Methods. 2021;18:945-52
130. Hurt RC, Buss MT, Duan M, Wong K, You MY, Sawyer DP. et al. Genomically mined acoustic reporter genes for real-time in vivo monitoring of tumors and tumor-homing bacteria. Nat Biotechnol. 2023;41:919-31
131. Airan R. Neuromodulation with nanoparticles. Science. 2017;357:465
132. Fan C-H, Wei K-C, Chiu N-H, Liao E-C, Wang H-C, Wu R-Y. et al. Sonogenetic-based neuromodulation for the amelioration of parkinson’s disease. Nano Lett. 2021;21:5967-76
133. Kalia LV, Lang AE. Parkinson’s disease. Lancet. 2015;386:896-912
134. Xian Q, Qiu Z, Murugappan S, Kala S, Wong KF, Li D. et al. Modulation of deep neural circuits with sonogenetics. Proc Natl Acad Sci U S A. 2023;120:e2220575120
135. Kanner AM, Bicchi MM. Antiseizure medications for adults with epilepsy: a review. JAMA. 2022;327:1269
136. Lane MA, Kahlenberg CA, Li Z, Kulandaival K, Secore KL, Thadani VM. et al. The implantation effect: delay in seizure occurrence with implantation of intracranial electrodes. Acta Neurol Scand. 2016;135:115-21
137. Muzumdar D, Patil M, Goel A, Ravat S, Sawant N, Shah U. Mesial temporal lobe epilepsy - an overview of surgical techniques. Int J Surg. 2016;36:411-9
138. Khazipov R. GABAergic synchronization in epilepsy. Cold Spring Harb Perspect Med. 2016;6:a022764
139. Phan T-N, Fan C-H, Wang H-C, Liu H-L, Lin Y-C, Yeh C-K. Modulation of GABAergic neurons in acute epilepsy using sonogenetics. J Control Release. 2025;377:341-53
140. Sjöstedt E, Zhong W, Fagerberg L, Karlsson M, Mitsios N, Adori C. et al. An atlas of the protein-coding genes in the human, pig, and mouse brain. Science. 2020;367:eaay5947
141. Kleinlogel S, Vogl C, Jeschke M, Neef J, Moser T. Emerging approaches for restoration of hearing and vision. Physiol Rev. 2020;100:1467-525
142. Sahel J-A, Boulanger-Scemama E, Pagot C, Arleo A, Galluppi F, Martel JN. et al. Partial recovery of visual function in a blind patient after optogenetic therapy. Nat Med. 2021;27:1223-9
143. Ji J, Gong C, Lu G, Zhang J, Liu B, Liu X. et al. Potential of ultrasound stimulation and sonogenetics in vision restoration: a narrative review. Neural Regen Res. 2024;20:3501-16
144. VanRullen R. Perceptual cycles. Trends Cogn Sci. 2016;20:723-35
145. Pinta CD la, Martín M, Sancho S. Rectal adenocarcinoma: clinical predictive factors for tumor response after chemoradiotherapy. Ann Oncol. 2019;30:iv22-3
146. Song Y, Chen J, Zhang C, Xin L, Li Q, Liu Y. et al. Mechanosensitive channel Piezo1 induces cell apoptosis in pancreatic cancer by ultrasound with microbubbles. iScience. 2022;25:103733
147. Steinbeck JA, Choi SJ, Mrejeru A, Ganat Y, Deisseroth K, Sulzer D. et al. Optogenetics enables functional analysis of human embryonic stem cell-derived grafts in a parkinson’s disease model. Nat Biotechnol. 2015;33:204-9
148. Qian X, Zheng Y, Chen Y. Micro/nanoparticle-augmented sonodynamic therapy (SDT): breaking the depth shallow of photoactivation. Adv Mater. 2016;28:8097-129
149. Hahmann J, Ishaqat A, Lammers T, Herrmann A. Sonogenetics for Monitoring and Modulating Biomolecular Function by Ultrasound. Angew Chem Int Ed. 2024;63:e202317112
150. Bansal A, Shikha S, Zhang Y. Towards translational optogenetics. Nat Biomed Eng. 2022;7:349-69
151. Choi J, Shin E, Lee J, Devarasou S, Kim D, Shin JH. et al. Light-stimulated insulin secretion from pancreatic islet-like organoids derived from human pluripotent stem cells. Mol Ther J Am Soc Gene Ther. 2023;31:1480-95
152. Latypova AA, Yaremenko AV, Pechnikova NA, Minin AS, Zubarev IV. Magnetogenetics as a promising tool for controlling cellular signaling pathways. J Nanobiotechnology. 2024;22:327
153. Stanley SA, Sauer J, Kane RS, Dordick JS, Friedman JM. Remote regulation of glucose homeostasis in mice using genetically encoded nanoparticles. Nat Med. 2015;21:92-8
154. Sawada T, Iino Y, Yoshida K, Okazaki H, Nomura S, Shimizu C. et al. Prefrontal synaptic regulation of homeostatic sleep pressure revealed through synaptic chemogenetics. Science. 2024;385:1459-65
155. Ferreira J. Using chemogenetics to reduce rat obesity. Lab Anim. 2024;53:172
156. Miyakawa N, Nagai Y, Hori Y, Mimura K, Orihara A, Oyama K. et al. Chemogenetic attenuation of cortical seizures in nonhuman primates. Nat Commun. 2023;14:971
157. Shin J, Kong C, Cho JS, Lee J, Koh CS, Yoon M-S. et al. Focused ultrasound-mediated noninvasive blood-brain barrier modulation: preclinical examination of efficacy and safety in various sonication parameters. Neurosurg Focus. 2018;44:E15
158. Guo H, Hamilton M, Offutt SJ, Gloeckner CD, Li T, Kim Y. et al. Ultrasound Produces Extensive Brain Activation via a Cochlear Pathway. Neuron. 2018;98:1020-1030.e4
159. Braun V, Blackmore J, Cleveland RO, Butler CR. Transcranial ultrasound stimulation in humans is associated with an auditory confound that can be effectively masked. Brain Stimul. 2020;13:1527-34
160. Wang W, Tang KWK, Pyatnitskiy I, Liu X, Shi X, Huo D. et al. Ultrasound-induced cascade amplification in a mechanoluminescent nanotransducer for enhanced sono-optogenetic deep brain stimulation. ACS Nano. 2023;17:24936-46
161. Chen W, Mehlkop O, Scharn A, Nolte H, Klemm P, Henschke S. et al. Nutrient-sensing AgRP neurons relay control of liver autophagy during energy deprivation. Cell Metab. 2023;35:786-806.e13
Author contact
![]() Corresponding author: Haitao Ran, Department of Ultrasound, the Second Affiliated Hospital of Chongqing Medical University, Chongqing, 400010, PR China. Email: ranhaitaoedu.cn.
Corresponding author: Haitao Ran, Department of Ultrasound, the Second Affiliated Hospital of Chongqing Medical University, Chongqing, 400010, PR China. Email: ranhaitaoedu.cn.