Impact Factor
- Issue 14; 2026
- Issue 13; 2026
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Volume 16; 2026
- Advance Articles
- Past Issues
- Cover Images
- Cover Suggestion
- Index & Coverage
- Special Issues
Introduction
Development and function of B...
B lymphocytes in cancer immunity...
Critical subsets of...
The crosstalk between TIBs and...
The prognostic and predictive...
Insights into mechanism research...
Prospects of B cells and TLS in...
Conclusions and perspectives
Abbreviations
Acknowledgements
References
International Journal of Biological Sciences
International Journal of Medical Sciences
 Global reach, higher impact
Global reach, higher impact

Theranostics 2025; 15(2):605-631. doi:10.7150/thno.105423 This issue Cite
Review
B cells and tertiary lymphoid structures in tumors: immunity cycle, clinical impact, and therapeutic applications
Xing Wu1,2, Qibo Huang1,2, Xiaoping Chen1,2, Binhao Zhang1,2 ![]() , Junnan Liang1,2
, Junnan Liang1,2 ![]() , Bixiang Zhang1,2
, Bixiang Zhang1,2 ![]()
1. Hepatic Surgery Center, and Hubei Province for the Clinical Medicine Research Center of Hepatic Surgery, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, Hubei, P. R. China.
2. Hubei Key Laboratory of Hepato-Pancreatic-Biliary Diseases, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, Hubei, P. R. China.
Received 2024-10-18; Accepted 2024-11-17; Published 2025-1-1
Abstract

Tumorigenesis involves a multifaceted and heterogeneous interplay characterized by perturbations in individual immune surveillance. Tumor-infiltrating lymphocytes, as orchestrators of adaptive immune responses, constitute the principal component of tumor immunity. Over the past decade, the functions of tumor-specific T cells have been extensively elucidated, whereas current understanding and research regarding intratumoral B cells remain inadequate and underexplored. The delineation of B cell subsets is contingent upon distinct surface proteins and the specific transcription factors that define these subsets have yet to be fully described. Consequently, there is a pressing need for extensive and comprehensive exploration into tumor-infiltrating B cells and their cancer biology. Notably, B cells and other cellular entities assemble within the tumor milieu to establish tertiary lymphoid structures that facilitate localized immune activation and furnish novel insights for tumor research. It is of great significance to develop therapeutic strategies based on B cells, antibodies, and tertiary lymphoid structures. In this review, we address the role of B cells and tertiary lymphoid structures in tumor microenvironment, with the highlight on their spatiotemporal effect, prognostic value and therapeutic applications in tumor immunity.
Keywords: tumor immunity, B cell, tertiary lymphoid structure, atypical memory B cell, immunotherapy
Introduction
Tumor immunology has provided new perspectives and methodologies for cancer treatment with deepening research on the tumor microenvironment (TME). The essence of tumor immunity lies in harnessing the human immune system to recognize, respond to, and eliminate tumor cells, thereby achieving effective immune clearance of the tumor. The tumor immune microenvironment stands as a pivotal focus in tumor immunology research, encompassing intricate interactions between tumor cells and immune cells [1]. Although the significance of T cells in cancer therapy and prognosis has been well established, a spectrum of strategies—from immune checkpoint blockade (ICB) therapy to adoptive T cell therapy—has revolutionized the paradigm of cancer treatment. B lymphocytes are typically less abundant and are not the spotlight in tumors, however, a multitude of recent data has indicated that tumor-specific B cells are relevant to favorable immunotherapy response and outcomes in patients.
Notably, tumor-infiltrating B cells (TIBs) are functionally diverse and heterogeneous. B cells and other immune cells are attracted to tumors and become progressively organized, evolving from the small lymphocyte aggregates into tertiary lymphoid structures (TLS). TIBs could engage in establishing immunosurveillance and potentiating antitumor immunity within the framework of tumor-associated TLS through cytokine secretion, antibody production, and antigen presentation. Nevertheless, the immunosuppressive TME, coupled with the structural disparities between assembled TLS and SLO, culminates in a compromised state of humoral immunity within TIBs and TLS. Leveraging engineered B cells, antibodies, and biocompatible TLS to reconfigure the TME may architect pioneering approaches in cancer therapy. Herein, we delve into the burgeoning roles of intratumoral B cells and TLS in immunity cycle, and illuminate their profound impact on patient prognosis and cancer treatment.
Development and function of B cells and tertiary lymphoid structures
B-cell development is coordinated by relevant transcription factors in specific locations and environments. Hematopoietic stem cells (HSCs) upregulate transcription factors such as Early B-cell factor 1 (EBF1), Forkhead box O1 (FOXO1), Paired box 5 (PAX5), and Interferon regulatory factor 4/8 (IRF4/8), leading to lineage commitment and the generation of common lymphoid progenitors (CLPs). Through the acquisition of CD19, CD20, and BCR, progenitor B cells (pro-B cells) develop into precursor B cells (pre-B cells), and eventually immature B cells [2], which are terminally activated in the periphery through T-cell dependent and T-cell independent manners. B cells undergo further proliferation and activation within SLO and TLS. Terminally differentiated plasma cells and memory B cells execute their function through antibody production to neutralize antigens and drive antibody-dependent cellular phagocytosis (ADCP), cellular cytotoxicity (ADCC) as well as complement-dependent cytotoxicity (CDC). Recent studies have indicated that B cells and antibodies play dual roles in tumor immunity.
Origin and differentiation of B cells in the bone marrow
B cells, including B1 and B2 cells, mediate the immune effects of antigen presentation, secretion of cytokines, and production of specific antibodies in infections, autoimmunity, and tumor diseases. CD5+ B1 cells, derived from fetal liver or hematopoietic stem cells, dominate the development of B cells in the late embryonic and early fetal stages, and underlie the immune responses of mucosa and epithelial tissue in the early stages of individuals. B cells are often referred to as CD5- B2 cells. B2 cells originate from HSCs and serve as important components of humoral immunity during late fetal development and after birth. B cells initially differentiate and develop into pro-B cells and begin to express Igα/Igβ heterodimers. Pre-B cells come to express a µ heavy chain and form the pre-BCR complex after the Pro-B cells complete the VDJ gene segment rearrangement. Through negative selection, they generate immature B cells with mlgM and gain central tolerance through clonal deletion, receptor editing, and anergy. Finally, immature B cells that start to express mlgD, gradually egress into the bone marrow and enter the blood circulation to become transitional B cells.
Immune response of B cells in the marginal zone, germinal center and extrafollicular region
In the peripheral lymphoid organs, transitional B cells further develop into mature B cells. It has been reported that 20% of circulating naïve B cells remain self-reactive and experience peripheral tolerance, fortunately some of which can reparticipate in immune responses through clonal redemption [3]. According to their distinct maturation sites, mature B cells are sorted into marginal zone B cells (MZ BCs), follicle-germinal center cells (GC BCs), and extrafollicular B cells (EF BCs). Finally, B cells are activated, proliferate, and differentiate into plasmablasts, plasma cells, and memory B cells. The activation of B cells involves two pathways: T-cell dependent and T-cell independent pathways in the peripheral lymphoid organs (Figure 1A).
B cell development and genesis of tertiary lymphoid structures. (A) The development and maturation of the B1 lineage and B2 lineage cells. (B) Formation, maturation and location of classic TLS in tumors. In the context of long-term inflammation or high tumor immunogenicity, tissue stromal cells or lymphocytes (LTo cells) produce numerous inflammatory factors (TNFα and IL-7) and chemokines (CXCL12/13, CCL19/21 and CCL5/22). The recruited LTi cells subsequently interact with stromal cells through the LTα1β2-LTβR axis, prompting the release of VEGFC and formation of HEV. In addition, released IL-17 promotes the release various chemokines of LTo cells, which recruit lymphocytes to express LTα1β2. The adhesion molecules secreted by vascular endothelial cells, as well as chemokines, further participate in the genesis of TLS by recruiting lymphocytes through HEV. The maturity, location, and density of TLS directly affect the efficiency of immune response. Based on the degree of maturity, TLS can be divided into early, primary follicular, and secondary follicular TLS. Early TLS are composed of dense lymphocyte aggregates and lack FDCs. Primary follicular TLS begin to appear with CD21+CD23- FDCs without GC formation, while TLS with high GC activity and CD21+CD23- FDCs are presented as secondary follicular TLS. TLS can be divided into intratumoral, peripheral infiltrating, and peritumoral based on their distribution within the tumor. TLS provide a local and vital microenvironment for the immune response in tumors. LTi: lymphoid tissue inducing cell; LTo: lymphoid tissue organizer cell; HEV: high endothelial vein; VEGFC: vascular endothelial growth factor C.
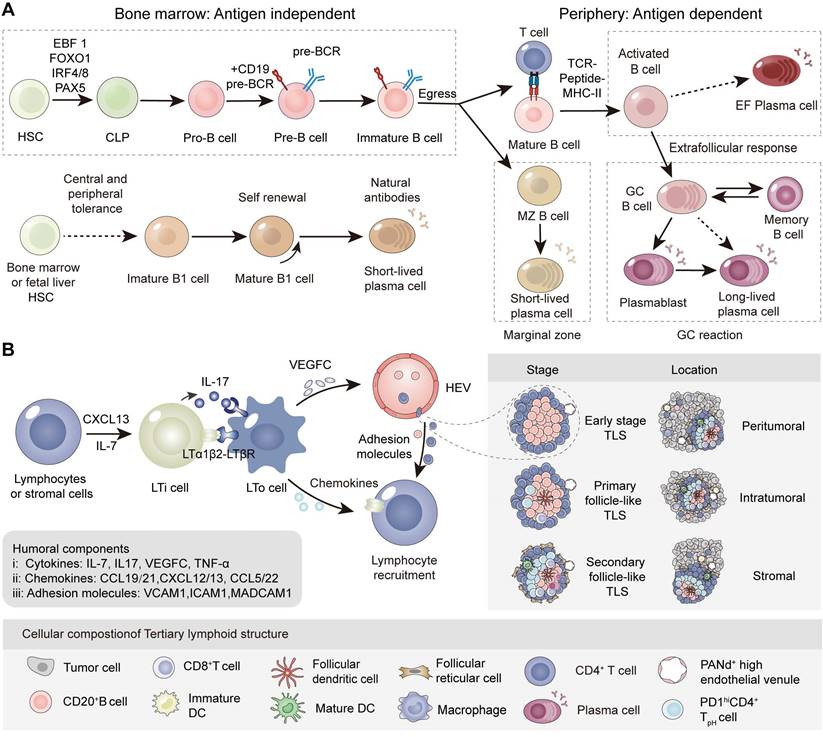
In the classical T-cell dependent pathway, B cells recognize the antigen presented by follicular dendritic cells (FDCs) through the BCR. T follicular helper (Tfh) cells stimulated by the same antigen interact with B cells at the edge of the T-cell zone and provide CD40L and cytokines for B-cell activation. Activation of the T-cell dependent pathway depends on interactions among FDCs Tfh and B cells. Ki67+CD23+ B cells undergo rapid proliferation in the dark zone of the germinal center accompanied by somatic hypermutation (SHM). Subsequently, B cells proceed to antibody class switching and affinity maturation in the bright zone, transforming from the secreting IgM type to IgA, IgE and IgG isotypes. This is essential for the different types of antigens reactions. Cytidine deaminase primarily expressed in activated B cells, is vital for the SHM and CSR processes and is involved in the formation of GC.
T-cell independent activation requires BCR to aggregate on the surface of B cells to cross-link with repeated antigen epitopes, while Toll-like receptor (TLR) stimulation provides a second activation signal. Activated B cells generate short-lived plasma cells that secrete IgM. T-cell independent pathways primarily occur in MZ BC and B1 BC (naïve-like B cells), which are important for rapid and early responses to drainage area pathogens such as blood and cavity pathogens [4].
B Cell mobilization center in the peripheral: SLO and TLS
Tertiary lymphoid structures, also known as tertiary lymphoid organs or ectopic lymphoid structures, are organized immune cell aggregates acquired from ectopic lymphoid tissues under non-physiological conditions [5]. TLS usually occur in chronic infectious tissues, autoimmune diseases and cancer. TLS have an organization similar to that of secondary lymphoid organs, including T cell zones, B cell follicles and high endothelial veins (HEV), but lack an encapsulated structure and afferent/efferent lymphatic vessels [6]. There are various reports on the formation of tumor-associated TLS, stemming from studies on ectopic lymphoid tissues or organs in tumors. The initial stage of TLS formation partially depends on the secretion of IL-7 and CXCL13 by stromal cells and lymphocytes at the site of inflammation or high tumor mutation burden (TMB) [7], supporting the recruitment of lymphoid tissue inducing cells (LTi cells) such as Th17 cells, B cells, macrophages and innate lymphoid cell-3 (ILC3s) [8]. Subsequently, LTi cells interact with lymphoid tissue organizer cells (LTo cells) via the LTα1β2/LTβR and IL-17/IL-17R axes, secreting a variety of effector molecules involved in peripheral node addressin-positive (PNAd+) HEV formation, lymphocyte recruitment (CXCL12/13, CCL19/21 and adhesion molecules) and ultimately TLS assembly [9]. Although the mechanism of tumor-associated TLS formation is still controversial, the main research is analogous to the formation of SLO. It has been demonstrated that the microbiota present within the TME influences the formation of TLS [10] when solid tumors develop on barrier surfaces including lung or colorectal tissues (Figure 1B) [11]. Intriguingly, recent reports have described a TLS with distinct architecture, wherein B cells are positioned at the periphery of the TLS.
Notably, the abundance, functional state, and distribution of B cells in TME are highly dependent on the location, density, and maturity of tertiary lymphoid structures [5], typically disseminated in the intratumoral and peritumoral sites. Intriguingly, immature TLS are more common than TLS with GC [12]. Studies have shown that B cells prefer to form a suppressive TME in immature TLS, while mature TLS in tumors usually serve as hubs of antitumor immunity [13]. These findings hold promise as a complementary approach through inducing the mature and high-density TLS in cancer treatment.
B lymphocytes in cancer immunity cycle: Intratumoral cycle and humoral circulation pathway
Current studies have concluded the immune effects of tumor-infiltrating T lymphocytes (TILs) within the cancer immunity cycle [14]. Our research group become intrigued by the source, tumor site, and distribution of tumor-infiltrating B cells. In the following section, we dissect the source of TIBs and distinguish them from TILs. Naïve B and naïve T cells within the tumor-associated TLS are generally thought to be derived from PNAd+ HEV [15], whereas the originally activated T lymphocytes and B lymphocytes in lymph nodes have the potential to migrate to the CD31+CD34+ tumoral vasculature. TIBs are either accumulated in the margin of tumors [16] or assembled from less organized lympho-myeloid aggregates (LMAs) to structured TLS. Next, we attempt to provide a detailed description of the spatial immune effects of the response patterns of TIBs (Table 1).
B cell activation pathways in cancer immunity cycle
| Feature | Humoral Circulation Activation Pathway | Intratumoral Cycle Activation Pathway |
|---|---|---|
| Activation Location | Secondary lymphoid organs | Tumor-induced TLS and LMAs |
| Activation& Response Path | Lymphatic and blood circulation | Recruitment from HEV; Response along fibroblastic tracks |
| Activation Manners | Lymph nodes: GC and EF; Spleen: marginal zone | GC pathway and EF pathway |
| Antigen Source | Circulating and draining tumor antigens | Intratumoral tumor antigens |
| Antigen Presentation | DCs, Macrophages | B cells offering the additional stimulus |
| Humoral immunity | B cells fully activated in peripheral lymphatic organs | Highly dependent on the distribution, density and maturity of tumor-associated TLS; Effective B cell clones fewer than TMB |
| Antibody Effect | Superficial layer of solid tumors, consistent with the vascular distribution | Inside the tumor derived from antibody secreting cells |
| Response Efficiency | Indirect, hysteretic but essential | Direct, efficient, but impaired |
| Relation | Initiation of intratumoral immunity | Extension and supplement of circulatory immunity trafficking into tumor core |
| Applications | Reconstitution of intratumoral immunity in the early postoperative period | In-situ response speed determining postoperative tumor status: tumor shrink, persistence, recurrence or metastasis |
Intratumoral cycle activation pathways based on tertiary lymphoid structures
The intratumoral immunity cycle of B cells is mainly based on TLS and lympho-myeloid aggregates. Owing to the notable differences in the therapeutic prognosis of these two pathways, we categorize the intratumoral immunity of B cells into GC pathway and EF pathway [17]. Activated T and B cells in these pathways eventually participate in the killing of tumor cells and accompanied by the release of tumor antigens, which can be regarded as the initiation of intratumor immunity.
The main function of TLS-GC is to produce memory B cells and long-lived plasma cells that secrete high affinity antibodies in tumors. In GC pathway, CXCL13+ Th cells in nasopharyngeal carcinoma (NPC) and breast cancer [18] are reported involving in the recruitment of GC B cells and CXCR5+ Tfh cells in tumor-associated TLS [19]. In addition to mature dendritic cells and macrophages, TIBs can function as antigen-presenting cells (APCs) by presenting antigens to T cells, thereby promoting the activation and proliferation of CD4+ and CD8+ T cells [20]. Furthermore, TLS associated follicular regulatory T cells (Tfr), regulatory B cells (Bregs), and regulatory T cells (Tregs) participate in the regulation of B and T cell activation by secreting multiple cytokines [10,21] (Figure 2B).
B cells in antitumor immunity: humoral circulation and local priming. (A) Schematic diagram of the B cells in cancer immunity cycle. (1) Intratumoral and circulating antibodies assist effector T cells in killing cancer cells through the immune response of neutralization, opsonization, ADCC, and CDC. (2) The release of tumor antigens or antigen-antibody complexes are taken up by APCs in tumors and are presented to lymphocytes in LMAs. (3) Next, APCs migrate to tumor-draining lymph nodes or TLS. (4) Naïve B cells in TDLNs are activated with the assistance of FDCs and CD4+ Tfh, and finally differentiate into affinity-matched plasma cells and memory B cells following SHM and CSR. (5) In addition, lymphocytes can migrate to tumor tissues through the circulation system under specific chemokines and adhesion molecules; antibodies secreted by plasma cells can diffuse throughout the body and bind to tumor tissue. (6) Lymphocytes infiltrate tumor tissues through HEV, tumor blood vessels and stroma, recognizing, secreting effector molecules, and killing tumor cells. (B) Intratumoral cycle activation pathway based on tertiary lymphoid structures. Compared with the GC pathway, naïve B cells are activated with the help of FDCs and CD4+ Tph in EF pathway and then gradually differentiate into atypical memory B cells and short-lived plasma cells. The short-lived plasma cells secrete mostly meaningless, low-affinity and self-reactive antibodies
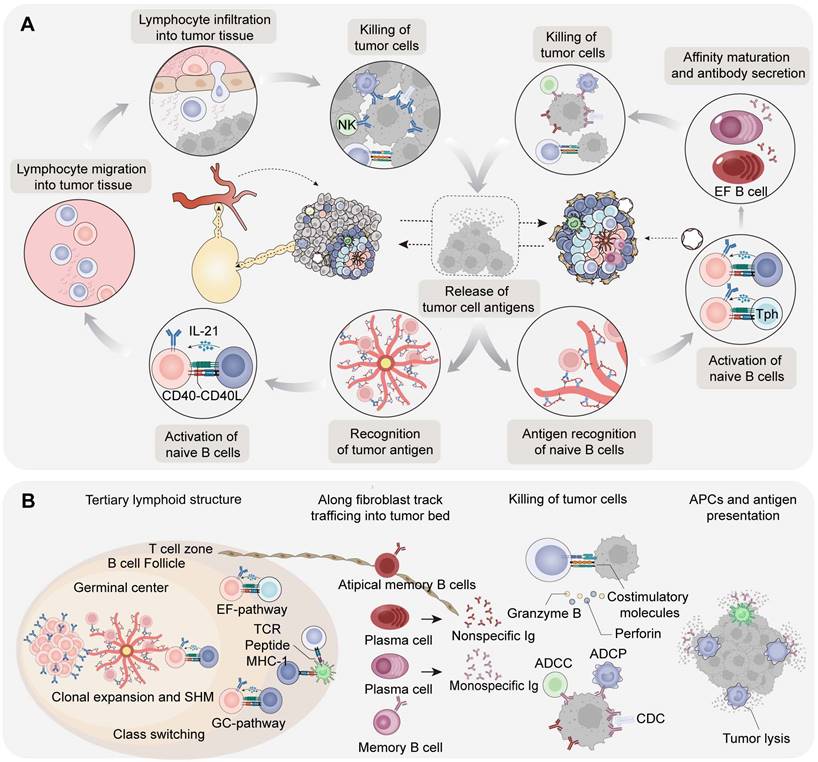
Recent studies have revealed that the extrafollicular (EF) pathway of B cells is independent of GC and enhanced in partial cancer [22]. The precursors of antibody-secreting cells (ASCs) in the EF pathway exhibit an atypical memory B-cell phenotype (AtM B cells). Further analysis revealed that the EF pathway is dominant in hepatocellular carcinoma (HCC), pancreatic adenocarcinoma (PDAC), and cervical squamous cell carcinoma. Both AtM B cells in the EF pathway and classical memory B cells in the GC pathway originate from naïve B cells, whereas T helper cells in the GC and EF pathways are distinct. T peripheral helper (Tph) cells in EF pathway are involved in inducing the formation of AtM B cells and show an exhaustive phenotype [23]. Naïve B cells mediate the activation of EF pathway B cells under the auxiliary stimulation of PD-1hiCXCL13+CXCR5- Tph from the T-cell area and tumor antigen complexes presented by FDCs. ASCs ultimately differentiate into AtM B cells and short-lived plasma cells which secrete low-affinity antibodies and begin to migrate away from TLS. In spatial TME architecture of renal cell carcinoma (RCC) patients, plasma cells generated in tumor-associated TLS were observed to further disseminate into the tumor bed along CXCL12+ fibroblastic tracks [24]. It can be speculated that the EF pathway and functional exhaustion of AtM B cells are upregulated in tumors, seen as an adaptive response or metabolic reprogramming.
Humoral circulation activation pathways based on secondary lymphoid organ
The humoral circulation activation pathway, especially in tumors that lack mature TLS, serves as another pathway of immunity cycle in B cells. Evidence suggests that B cells engage in a systemic response to tumor antigens, as indicated by tumor-specific B cells, plasma cells and antibodies detected in the peripheral blood [25]. Based on previously reported tumor antigens found in the serum, tumor-specific B cells in peripheral lymphatic organs may be selectively activated and produce antibodies when tumor antigens are released into the blood or presented by APCs through lymphatic vessels [26]. These steps allow lymphocytes to proliferate and differentiate into effector T cells and memory B cells, which migrate into the tumor bed and lead to the destruction of tumor cells [27]. It has been hypothesized that activated plasma cells may even migrate to the bone marrow for long-term survival and antibody production under chronic stimuli. The antibody continuously secreted by plasma cells in the SLO will eventually spread to the tumor bed through lymphatic and blood circulation.
In particular, B cells in peripheral lymphoid tissue and circulation may be attracted directly to the tumor bed instead of exclusively through HEV of TLS, despite the fact that they are less abundant in tumors without TLS [28,29]. In TLS-deficient tumors, it has been postulated that B cells can be recruited through conventional CD31+CD34+ blood vessels [21]. Tumor-infiltrating immune cells and fibroblasts in tumor tissues could secrete chemokines such as CXCL13 to produce chemotactic effects on B cell subsets. In addition, CXCL9 and CXCL10 secreted by the intestinal epithelium can participate in the recruitment of IgA+ plasma cells in mucosal tumor such as colorectal cancer [30]. Interestingly, owing to the lack of effective vascular systems in solid tumors, circulating antibodies and immune cells may only penetrate the superficial layer of the tumor, which is consistent with the vascular distribution of the tumor [31]. Therefore, TLS-based intratumoral immunity may be a strategy for the penetration of immune cells to antagonize the nutrient-poor and the oxygen-deficient tumor cores (Figure 2A).
Critical subsets of tumor-Infiltrating B cells in TME
Subsets of TIBs and antibodies have attracted increasing attention in cancer research and clinical practice. Several studies have focused on profiling the immune landscape of B cells and plasma cells across distinct cancers through sequencing technology [22,32]. The imbalanced status is revealed in intratumoral regulatory B cells and functional B cells including the components of antitumor plasma cells and memory B cells, induced by immunosuppressive TME.
Regulatory B cells in TME
The role of regulatory B cells has been widely reported in infections, autoimmune diseases, transplantation, allergic diseases and tumors. Bregs, originally identified in autoimmune enteritis, are a subset with immunosuppressive function that play an important role in individual immune homeostasis [33]. However, the specific transcription factors associated with Bregs have not been clearly characterized, posing challenges for research in this field. It still remains controversial whether Breg constitute an independent subset or a regulatory B-cell state induced under distinct conditions. Supporting this view, it is currently found that immature B cells [34,35], plasma cells [30], plasmablasts [36] and even B1 cells [37,38] all have the potential to differentiate into Bregs and produce IL-10. In this section, we introduce the cytokines and metabolites involved in Breg induction, intracellular biological pathways and Breg effector functions in tumors (Figure 3).
Functional B cells and regulatory B cells in the tumor microenvironment. Components in TME could participate in tumor-induced regulatory B cells through epigenetic modifications, transcriptome regulation, and metabolic reprogramming. HA, LTB4, lactate, and exosomes can promote transfer of B cells into regulatory phenotypes by acting as BCR agonists, TLR agonists, CD40 agonists or Sting agonists. Bregs can exert regulatory functions by upregulating impressive surface markers and the secretion of multiple inhibitory cytokines. ROS: reactive oxygen species; PlGF: placenta growth factor; L-kyn: L-kynurenine; LTB4: leukotrienes B4; HA: hyaluronic acid; HMGB1: high mobility group box 1; AHR: aryl hydrocarbon receptor; PPARα: peroxisome proliferator-activated receptor α; TIM: T-cell immunoglobulin and mucin-domain; MDSC: myeloid-derived suppressive cell; LT: lymphotoxin; TRAIL: TNF-related apoptosis-inducing ligand; GABA: γ-Aminobutyric Acid; ADO: adenosine; GrB granzyme B; IDO: indoleamine-2,3-dioxygenase; 5-LO: 5-lipoxygenase.
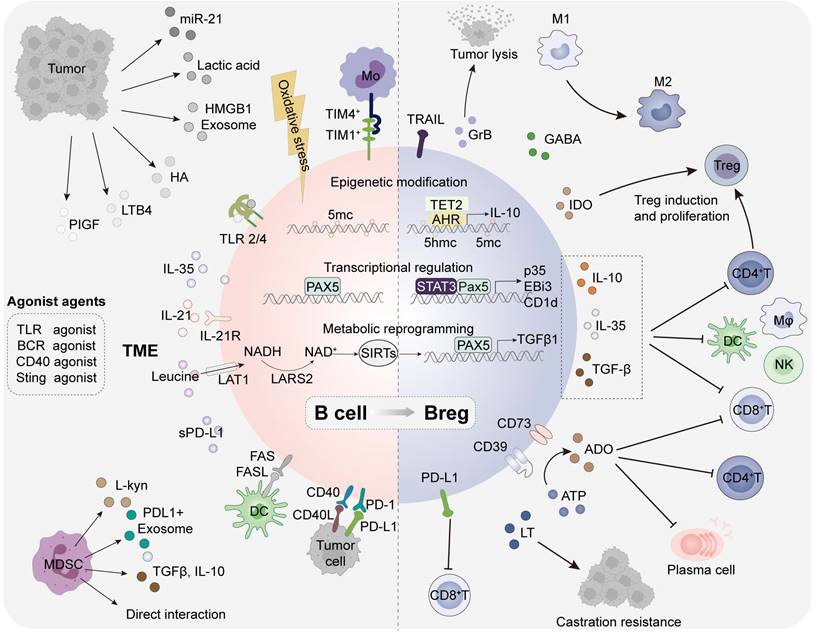
Indeed, Bregs with multiple surface markers have been identified in human and mouse tumors, peripheral blood, and draining lymph nodes (Table 2). TME derived soluble molecules (IL-1β [39], IL-21 [40,41], IL-35 [42], ROS [43], PlGF [44], miR-21 [45], and sPD-L1 [46]), metabolites (Leucine [47], L-kyu [48], LTB4 [49], hyaluronan [50], and lactic acid), exosomes [51] (PD-L1 [52] and HMBG1 [53]) and cell-interactions have been reported as involving in the induction of Bregs. Given that most Bregs are induced at different B cell stages, it is particularly critical to explore and evaluate the environmental factors that drive Breg production. These molecules can promote Breg differentiation by binding to specific receptors (IL-21R [40], AHR [48], PPARα [49], TLR2/4 [53] and BCR [54]) of B cells. Intriguingly, many receptor agonists (BCR agonists [55], TLR agonists [50,53] CD40 agonists [56] and STING agonists [57]) have been unveiled in B cell activation but may also trigger Breg differentiation in some cases, possibly related to the stimulation duration and concentration of these agents. In addition, multiple contact-dependent mechanisms have been uncovered in Breg generation, including CD40/CD40L and PD-1/PD-L1 by tumor cells, TIM-1/TIM-4 by CD11b+ myeloid cells, CD95/CD95L by semimature DCs, PD1/PDL1 by MDSCs.
Regulatory B cells identified in human and murine cancers
| Cancer type | Surface marker | Location | Effector function | Refs |
|---|---|---|---|---|
| HNSCC | CD19+ CD19+CD73+CD39+CD23+CD21- | Human; PBMC Human; PBMC, Tumor | IL-10, PD-1 Adenosine | [51] [138] |
| Breast carcer | CD19+ CD24hiCD27+ | Human; Tumor Human; TDLN | Granzyme B IL-6, TNF-α | [41] [143] |
| Gastric cancer | CD19+CD24hiCD38hi CD19+CD24hiCD27+ | Human; PBMC, Tumor Human; PBMC, Tumor | IL-10, TGF-β1 IL-10 | [34] [157] |
| HCC | CD19+CD5hi CD27hiCD38dim CD5hiCD24-CD27-/+CD38+/hi | Human; Tumor Human; PBMC, Tumor | PD-1, IL-10 IL-10 | [50] [53] |
| PDAC | CD19+CD24hiCD38hi CD38+CD19+ | Human; PBMC Human; Tumor | IL-35, IL-10 IL-10 | [35] [166] |
| CRC | CD19loCD27hi CD19+B220-CD20-IgD-CD38+CD27+ CD138- | Human; Tumor Human; Tumor | IL-10 TGF-β1 | [36] [47] |
| Ovarian carcinoma | CD19+CD20+ CD19+ | Human; Ascites Human; Tumor | IL-10 Granzyme B | [172] [41] |
| Cutaneous squamous cell carcinoma | CD19+CD73-CD25+CD71+TIM1+CD154+ | Human; PBMC | IL-10 | [56] |
| Breast cancer | CD19+CD81hiCD25+ | Murine; Tumor | TGF-β1 | [49] |
| HCC | CD19+ CD19lowIgA+B220-CD138+ MHC-IIlow | Murine; Tumor Murine; Tumor | PD-L1, IL-10, Fas-L PD-L1, IL-10, TGF-β1 | [40] [58] |
| PDAC | CD19+CD1dhiCD5+ CD19+CD1dhiCD5+CD21hi CD19+PD-L1+ | Murine; Tumor Murine; Tumor Murine; Tumor | IL-35 IL-35 PD-L1 | [37] [35,116] [39] |
| CRC | CD19+IgA+ | Murine; Tumor | PD-L1, IL-10, TGF-β1 | [30] |
| Prostate cancer | CD19+CD20loB220loIgA+PD-L1+ | Murine; Tumor | IL-10 | [68] |
| Melanoma | CD19+CD5+CD1dintCD43+ | Murine; Tumor | IL-10 | [38] |
TDLN: tumor-draining lymph nodes; PBMC: peripheral blood mononuclear cell
The total external stimulation triggers B cell intracellular epigenetic modification, transcriptional regulation and metabolic reprogramming, mediating Breg transformation. Triggered by oxidative stress in the HCC microenvironment, a predominant epigenetic regulatory enzyme ten-eleven translocation-2 (TET2) could promote the generation of Bregs by catalyzing the formation of 5-hydroxymethylcytosine (5-hmc) and enhance IL-10 transcription in B cells [43]. The dysregulated transcriptional program impeding plasma cells differentiation in PDAC involves an IL-35-induced STAT3-PAX5 signaling pathway that upregulates BCL6, promoting the transformation of naive B cells into Bregs and contributing to immune evasion [42]. In addition, leucine-rich diets associated leucine-tRNA-synthetase-2-expressing (LARS2+) Bregs in colorectal cancer (CRC) can lead to the TGF-β1 production and immunosuppression through mitochondrial NAD+ regeneration and oxidative metabolism [47].
Tumor induced Bregs resemble Tregs can participate in the formation of an immunosuppressive TME through the secretion of various effector molecules and the upregulation of surface markers [58]. The suppressive patterns are mainly divided into three types: conventional effector molecules (IL-10, IL-35, TGFβ, LT, PD-L1, FasL and TRAIL), plasma cell-related effector molecules (IgA and IgG), neurotransmitters and metabolites (GABA, ADO, GrB and IDO) [41]. They can inhibit the function of anti-tumor immune T cells and natural killer (NK) cells, promote the generation of Tregs, and suppress the effects of APCs through various mechanisms to potentiate tumor growth, proliferation, and immune evasion [36]. Bregs have been implicated in poor prognosis and drug resistance across various tumor types. The impact of TIBs can be either anti-tumorigenic or pro-tumorigenic, contingent on the specific malignancy and the equilibrium between functional B cells and regulatory B cells. A more detailed characterization of Breg subsets may offer significant advancements for cancer research.
Plasma cells and antibodies in TME
Tumor-infiltrating plasma cells are a group of highly heterogeneous cells and humoral immunity status dominates in distinct anti-tumor response. In poorly immunogenic tumors, plasma cells and antibodies form immune complexes that serve as an amplification loop and downregulate the antigen threshold required for T cell activation, enhancing T cell responses against tumor cells [59].
Although tumor-specific plasma cells selectively expand under the stimulation by tumor antigens, the effective clonal expansion is far less than the number of potential tumor neoantigens [60]. The contradictory situation of mutation-generated tumor antigens and effectively responsive B cells reflects impaired humoral immunity in tumors, which may be partially attributed to the suppressive microenvironment or the difference between tumor-induced TLS and well-structured lymph nodes. Tumor-associated antigens (TAAs) are usually described as highly expressed self-antigens including cancer germline antigens, while tumor-specific antigens (TSAs) are classified as genomic mutations and posttranscriptional aberrations of normal proteins. Mechanistically, this may also elucidate the coexistence that self-reactive plasma cells and auto-antibodies have been found in tumor tissues and peripheral blood [61], rendering either anti-tumor or pro-tumoral humoral responses. Coincidentally, two types of related growth factor receptor auto-antibodies isolated from B cells show opposite effects, with agonist antibodies promoting the growth of breast cancer cells and antagonizing the opposite of that in breast cancer patients [62]. In addition to antibodies targeting tumor neoantigens, there are also specific antibodies against microbial proteins in the context of viral infection-related tumors [63].
Intratumoral TLS are sites for the generation of mature B cell immunity, where plasma cells can migrate into the tumor bed along the fibroblastic tracks and secrete IgG or IgA antibodies [24]. IgG+ plasma cells, which are typically associated with improved clinical outcomes are preferentially present in inflamed tumors, while IgA+ plasma cells are predominant in mucosal epithelial tumors [64]. To be specific, it is found that glucose deprivation may suppress IgG+ plasma cell generation and antitumor immunity through the SATB1/STAT6 axis in colorectal cancer [65]. In addition, CXCL10-induced macrophages and IgG+ plasma cells contribute to the immunosuppressive microenvironment in HCC [66]. Furthermore, IgG+ plasma cells targeting heat shock protein 4 (HSPA4) in tumor-draining lymph nodes (TDLNs) participate in the formation of the premetastatic niche in breast cancer [67]. In contrast, IgA+ plasma cells expressing IL-10 or programmed death ligand 1 (PD-L1) dismantle the antitumor immunity of T cells in HCC and prostate cancer, whereas they are involved in the protective humoral response in ovarian cancer [58,60,68].
Memory B cells in TME
Human memory B cells, defined as classical class-switched CD21+CD27+ B cells and CD21-CD27- atypical memory B cells, are the reserve force of the immune responses and are more abundant than naïve B cells in the peripheral blood, especially in elderly individuals [69]. On the basis of the surface markers IgD and CD27, IgD-CD27- refers to double-negative B cells (DNBs). This section primarily introduces AtM B cells in tumors owing to the space limitations.
AtM B cells, characterized as CD11c+CD21-CD27-IgD-Tbet+ in human, are upregulated during aging, chronic infections, and autoimmune diseases and accumulate more significantly in female. The transcription factors T-bet and Zeb2 may be vital components of AtM B cells to achieve antibody class switching and antigen presentation [70]. They express immunosuppressive FCRL4, cytokine IL-10 and receptors, typically participating in the inhibition of anti-tumor response. As mentioned earlier, regardless of whether glutamine metabolic derivatives or Tph interacts, the transcription factor Zeb2 may be necessary for the formation of AtM B cells in tumors. Zeb2 can mediate the transformation of GC B cells to EF AtM B cells by downregulating Mef2b expression, thereby mediating the exhaustion of phenotypic B cells in tumors. An in-depth exploration of the molecular mechanism through which Zeb2 promotes the occurrence of AtM B cells, may present alternative ideas and outlooks on cancer research.
A single-cell B cell profiling reveals stress-responsive memory B cells and tumor-associated atypical B cells, two tumor-rich subsets with prognostic potential that are shared in a pan-cancerous manner. In particular, tumor-associated AtM B cells are characterized by high levels of clonal expansion and proliferation, as well as close interactions with activated CD4+ T cells in tumors, which can predict the immunotherapy response [32]. Another tumor-infiltrating B cell sequencing study demonstrated that atypical memory B cells developed by EF pathways, are independent of TLS-GC B cells and enriched in tumors, which are associated with shorter overall survival (OS) in patients [22]. An immunosuppressive AtM B cell subset was identified in a mouse model of bladder cancer, and depletion of this subset delays cancer progression, indicating that AtM B cells promote bladder cancer progression [71]. AtM B cells exhibit an exhausted phenotype and are increased in non-small cell lung cancer (NSCLC) and mesothelioma patients with non-response to ICB therapy, suggesting that AtM B cells may impede the ICB treatment response [72]. In addition, AtM B cells are enriched in the peripheral blood of patients with breast cancer compared with healthy controls, and their role needs to be further elucidated [73].
The crosstalk between TIBs and other tumor infiltrating immune cells
The interaction between B cells and T cells in TME
TIBs engage in intricate interactions with resident immune cells and cancer cells, which affects tumorigenesis and progression. The presence of TIBs close to CD8+ T cells or the co-localization of B cells and T cells in the intraepithelial infiltration of breast cancer, malignant melanoma and ovarian cancer are positive prognostic markers [74]. The close contact between TILs and TIBs indicates a functional interaction among them, which is related to enhanced local immune activation and helps to improve the prognosis of patients [75]. However, the coexistence of Bregs and Tregs in TIL aggregates is correlated with shorter metastasis-free survival in cancer patients [76]. Cho et al. indicated that impaired Tfh-B-Trm (tissue-resident memory T cell) cooperation in the formation of TLS, accompanied by dysregulated Trm homeostasis and loss of Tfh-B crosstalk, underlies the unfavorable anti-PD-1 response in EGFR-mutated lung cancer [77]. In a breast cancer mouse model treated with low-dose cyclophosphamide combined with CSF1R inhibitors, CD4+ T cells and antigen-presenting B cells were enriched and colocalized in the TLS, inducing the durable tumor regression after combination treatment [78].
Specifically, three types of CD4+ TIL responses to TIBs have been identified: activated, antigen-related and unresponsive TIBs. In the activated antigen-related CD4+ TIL population, activated TIBs are associated with the effector T cell response. Alternatively, exhausted TIBs are associated with the regulatory T cell phenotype [79], suggesting that targeting TIBs can serve as the potential therapeutic strategy for immunotherapy in NSCLC. Tumor neoantigens recognized by TILs promote the crosstalk between tumor-specific CD4+ T cells and tumor-specific B cells, thus enhancing the anti-tumor immunity of CD8+ T cells in lung adenocarcinoma (LUAD) [80]. Recent preclinical studies have shown that the generation of IL35+ B cells induced by STING agonists may attenuate the proliferation of NK cells and weaken the NK-driven anti-tumor response. It is suggested that the combination of inflammation-inducing STING agonists and IL-35 blockers may optimize this outcome [57].
The interaction between B cells and myeloid cells in TME
The crosstalk between tumor-infiltrating B cells and myeloid cells (macrophages, DCs, MDSCs, and neutrophils) has also been reported. Apart from direct cell-cell contact, soluble metabolites derived from B cells can perform immunomodulatory functions, which may be drug targets of the immune responses. B cells and plasma cells can secrete metabolites GABA, promoting the differentiation of monocytes into anti-inflammatory macrophages, which can secret IL-10 and inhibit the killing function of CD8+ T cells [81]. Besides, during cancer treatment, the macrophage layer of the subcapsular sinus in TDLNs is undermined, allowing tumor-derived extracellular vesicles to interact with B cells of lymph node cortex and mediate cancer-enhancing immunity [82]. IL17+ Th cells selectively recruit CXCR3+ B cells which can induce tumor-promoting macrophage polarization and facilitate hepatocellular carcinoma development [83].
Moreover, immune complexes deposited in precancerous tissue, promote squamous cell carcinogenesis by activating bone myeloid cells dependent on Fcγ receptors. More than 50% of B cells display an FcγRIIlow/- activation phenotype in HCC, and this phenotype is positively correlated with cancer progression. Further studies have revealed that semimature dendritic cells in tumors participate in the formation of FcγRIIlow/- B cell dependent on the CD95L pathway. FcγRIIlow/- B cells can secrete IL-10 to inhibit the anti-tumor response of cytotoxic T cells [84]. MDSCs, accumulated around the GC in the spleen of tumor-bearing mice, could promote B-cell production of IgA in a TNFR2-dependent manner [85]. Tumor-induced Bregs (tBregs) can initiate the regulatory subset functions of MDSCs and suppress antitumor immunity, which is partially dependent on TGFβR1/TGFβR2 signaling [86]. There is evidence that tumor-associated neutrophils drive B cell recruitment through the release of TNF-α and regulate plasma cell differentiation through neutrophil membranal B-cell activating factor (BAFF) [87].
The prognostic and predictive value of B cells and TLS in human cancer
The genetic traits of cancers, comprising tumor mutational burden, immune checkpoint expression and immune cell infiltration, are critical indicators of response to immunotherapy in patients with tumors [88]. The manifestation of TLS, along with intratumoral B cells, has been reported supporting the favorable outcomes and ICB response in both immunogenic tumors and TMB-low tumors [10]. Investigating the composition of tumor-associated TLS and TIBs can shed light on their contribution to the prediction and prognosis of cancer patients, supporting therapeutic approaches focused on B cells and TLS.
Methods for TIBs and TLS detection and evaluation in tumors
To fully characterize TIBs and TLS in tumors, it is critical to employ proper techniques to identify and analyze their signatures in the TME [27]. B cell detection and quantification can be achieved through methods such as flow cytometry, transcriptomic sequencing profiles, and immuno-imaging techniques. Conventional methods for detecting TLS rely on surgically obtained tumor tissue samples and include hematoxylin and eosin (H&E) staining, immunofluorescence (IF), multiplex immunohistochemistry (mIHC), RNA sequencing (RNA-seq), and spatial transcriptomics (ST) [89]. In addition, histological and genomic analyses can be applied to quantitatively evaluate TLS. Compared with the histological analysis, integrating multi-omics approaches may be further put into use to effectively and complementarily assess TLS in the future. The 12-chemokine signature (12-CS) has been identified to characterize the gene expression profiles of TLS [90], which connects a specific set of chemokine receptor/ligand genes with the formation and maintenance of TLS in tumors. The application of nanomaterials and artificial intelligence has driven the development of non-invasive imaging based on nanoprobes, such as CT and MRI [91]. These non-invasive imaging techniques provide new insights into the predictive and prognostic value of TLS [92].
The prognostic value of TIBs and TLS in patient survival
Previous reports have pointed out that the heterogeneity of tumor-infiltrating B cell subsets determines their dual roles of pro-tumor and anti-tumor. In preclinical studies, the use of CD20 antibodies to deplete tumor-infiltrating B cells displayed different effects on tumor progression. In fact, the prognostic study or depletion of the entire population of tumor-infiltrating B cells in pan-cancer can be compared to that of T cells. This suggests the need to conduct an in-depth exploration of the role of B cell subsets in tumors. Tumor-infiltrating B cells have been found to be associated with a better prognosis in NPC [93], head and neck squamous cell carcinoma (HNSCC) [94,95], breast cancer [96], gastric cancer [97], HCC, CRC [98], ovarian cancer [99], prostate cancer [100], and melanoma [101] (Table 3). This manifests as improved prognostic indicators such as OS, Disease-Free Survival (DFS), Disease-Specific Survival (DSS), and reduced risk of recurrence. However, high densities of CD8+ T cells and DC-LAMP+ DC cells are correlated with shorter OS in RCC patients and lung metastasis [101,102]. In lung adenocarcinoma patients with a high plasma cell gene score, there is less infiltration of B cells, CD8+ T cells, CD4+ T cells, and dendritic cells, indicating a weak immune response and a poor prognosis. Notably, high density infiltration and positive prognostic effects of B cells (except for Breg) may be highly dependent on mature TLS formation [24].
Depicting the prognostic and predictive value of TIBs and TLS in human cancers
| Tumor type | Analysis factor | Detecting method | Prognostic Indication | Prognostic value | Predictive value | Refs |
|---|---|---|---|---|---|---|
| GBM | TIBs, density | IF | Patient survival | Neutral | NA | [131] |
| NPC | TIBs | RNA-seq | PFS | Favorable | NA | [93] |
| HNSCC | TLS TLS, TIBs TLS, location | RNA-seq, IF RNA-seq, IF Multi-omics | RFS PFS, OS, RFS NA | Favorable Favorable NA | NA NA Positive | [103] [95] [117] |
| Lung cancer | TLS Plasma cells Plasma cells | H&E, IHC RNA-seq RNA-seq | PFS, OS OS OS | NA NA Adverse | Positive Positive NA | [147] [118] [145] |
| Breast cancer | TIBs, density TLS TLS, density | RNA-seq 12-CS H&E, IHC | DFS, OS PCR OS, PFS | Favorable NA Favorable | NA Positive NA | [96] [124] [104] |
| Gastric cancer | TLS, mature TLS, Trm TIBs | H&E IF IHC | OS OS, DFS OS, DFS | Favorable NA Favorable | NA Positive NA | [105] [120] [97] |
| HCC | TLS Adjacent tumoral TLS | H&E H&E | RFS, OS Early/Late recurrence | Favorable Neutral | NA NA | [106] [112] |
| PDAC | TLS, mature TLS, mature | RNA-seq, IF H&E, IHC | Survival probability OS, PFS | NA Favorable | Positive NA | [163] [107] |
| CRC | TLS TLS, location, density TLS, plasma cells | IHC, H&E IHC H&E, IHC, 12-CS | OS OS, RFS RFS, OS | Favorable Favorable Favorable | NA NA NA | [108] [111] [98] |
| ccRCC | TLS, TIBs, IgG TLS | ST IHC | PFS OS | NA Adverse | Positive NA | [24] [102] |
| Urothelial carcinoma | TLS TLS | IHC, H&E IHC | DFS OS, RFS | Favorable NA | NA Positive | [109] [125] |
| Prostate cancer | Plasma cells | IHC | MFS, DFS | Favorable | NA | [100] |
| Ovarian cancer | Plasma cells | RNA-seq | OS | Favorable | NA | [99] |
| Melanoma | TLS, TIBs, density TLS, TIBs | IF, RNA-seq H&E, IHC | OS OS | NA NA | Positive Positive | [101] [176] |
| Soft tissue sarcoma | TLS, TIBs TLS | IF, RNA-seq IHC | OS, PFS, ORR OS, PFS | NA NA | Positive Positive | [29] [126] |
IGKC: immunoglobulin kappa C; RFS: recurrence-free survival; FFS: failure-free survival; PFS: progression-free survival; MFS: metastasis-free survival.
The impact of TLS on the control of tumor development and metastasis has been widely reported. In HNSCC [103], lung cancer, breast cancer [104], gastric cancer [105], HCC [106], pancreatic cancer [107], and CRC [108], urothelial carcinoma [109], the TLS signature is associated with prolonged OS and DFS in patients. These results may be partly attributed to the fact that HEV and TLS facilitate the infiltration of circulating immune cells into tumors and initiate local immune responses. Indeed, HEV, 12-CS, and DC-LAMP, which serve as components of TLS, are frequently used as alternative indicators of tumor prognosis and have positive effects on a variety of tumors [110]. A multicenter propensity score-matched study indicated that patients with a high TLS density in right-sided colon cancer have a better prognosis than those with a low TLS density [108]. Another relevant study revealed that the distribution and abundance of TLS were associated with molecular subtypes and clinical outcomes in patients with colorectal cancer liver metastases but not in those with pulmonary metastases [111]. Additionally, mature TLS are believed to be associated with a reduced risk of early recurrence in hepatocellular carcinoma [112]. The location of TLS also shows differences in tumor prognosis, with TLS located farther away from the tumor being associated with poor prognosis, whereas TLS around the tumor predicts a favorable prognosis [113]. Nevertheless, TLS suppressive components, including TLS-associated Tregs and tumor cells detected are often associated with tumor progression and advanced stages [114]. Overall, one plausible explanation of the results is that TLS and TIBs can provide sites for T cell activation and additional antigen stimulation, which is beneficial for anti-tumor immunity and patient prognosis.
The predictive value of TIBs and TLS in response to cancer immunotherapy
Recent studies have shown that TIBs and TLS exert beneficial effects in predicting the clinical outcomes of cancer patients undergoing immunotherapy. It should be noted that on the one hand, TLS can serve as a predictor of the ICB response in tumor patients. On the other hand, neoadjuvant ICB treatment, radiotherapy, and chemotherapy can further induce TLS formation.
Lin et al. showed that CXCL13 secreted by tumor-infiltrating immune cells is significantly correlated with the immune responses to cancer immunotherapy in HNSCC [115]. However, IL-35 was reported as a negative indicator in antitumor immunity and response to immunotherapy in PDAC [116]. TLS-enriched type has a better prognosis and response to ICB therapy. The proximity of TLSs to tumor was found to be a key indicator of ICB response in HNSCC, while patients with TLS located far from tumor cells had a poorer prognosis [117]. Similar results were observed in lung squamous cell carcinoma. In advanced NSCLC, TLS has been shown to predict the response to ICB independently of PD-L1 expression and CD8+ T cell density, but not the outcome of chemotherapy [118]. In contrast to NSCLC, the density of follicular helper T cells in high-grade serous ovarian cancer (HGSOC) is not adequate enough to initiate functional B cell and mature TLS formation, which may be insufficient to maintain the ICB-sensitive TCF1+PD-1+CD8+ T cell phenotype [119]. In addition, the presence of TLS and Trm cells in tissues is conducive to a better response in gastric cancer patients undergoing anti-PD-1 therapy [120].
In human LUAD, tumor-binding antibodies against endogenous retroviruses (ERV) exert antitumor activity and ERV expression predicts the outcome of ICB therapy [121]. In patients with advanced TNBC responding to paclitaxel or its combination with the anti-PD-L1 atezolizumab, researchers found that follicular B cells and conventional type 1 DCs are considered as the TLS organizers and consistently increased after combination therapy [122]. In another clinical study on the spatial predictors of the immunotherapy response to TNBC, immune interactions between B cells and granzyme B+T cells were shown to be secondary predictive molecules after CD8+TCF+T cells [123]. A recent letter described the ability of TLS to predict the response to neoadjuvant chemotherapy and benefit from immunotherapy in HER2-negative breast cancer [124]. Helmink et al. assessed the favorable role of TIBs and TLS in response to immunotherapy with ICB treatment through bulk and single-cell RNA sequencing in patients with melanoma and RCC [101]. Similarly, in patients with urothelial carcinoma, high densities of B cells, T cells and TLS could predict the response to combined PD-1 and CTLA4 blockade treatment [125]. In a phase 2 clinical trial of soft-tissue sarcoma, B cells remained the strongest prognostic factor, even in the context of low CD8+ T cells and cytotoxic content. The class E, which is abundant in TLS and TIBs, has shown improved prognosis and a high response to PD-1 blockade with pembrolizumab [29]. Italiano et al. have revealed that the presence of TLS in advanced soft-tissue sarcoma is a potential predictive biomarker for improved patient response to pembrolizumab treatment [126]. These favorable results of immunotherapy are closely related to the high expression of inhibitory molecules in the TME and the functions of TLS and TIBs in tumors.
Insights into mechanism research of B cell in pan-cancer
Glioblastoma
Glioblastoma (GBM) is the most common malignant primary brain tumor in adults. ICB therapy has revolutionized cancer treatment; however, GBM patients have yet to benefit from this breakthrough. The primary obstacle in finding effective immunotherapy strategies is partially attributed to the low infiltration rate of TILs and a shortage of appropriate lymphatic vessels within the brain parenchyma [127]. A study of tumor infiltrating immune cell profiles revealed the lymphocyte proportion in GBM, primarily including CD4+ T (0.5%±0.7%) and CD8+ T cells (0.6% ± 0.7%), and smaller numbers of CD4-CD8-T cells (0.2% ± 0.4%), Tregs (0.1% ± 0.2%), B lymphocytes (0.1% ± 0.2%), and NK cells (0.05% ± 0.05 %) [128]. B cells and T cells are typically clustered within a radius of 15μm, while this proximity leads to the formation of immune synapses within the TME of GBM, mediating the inhibition of CD8+ T cell activation [129]. In addition, GBM-associated MDSCs could promote the regulatory function of B cells through delivering membrane-bound PD-L1, endowing Bregs with the potential to inhibit CD8+ T cell activation and acquisition of the effector function [52]. It has been reported that glioma cell-derived placental growth factor (PlGF) is also involved in the generation of regulatory B cells [44]. Another study revealed that the TGFβ-1/TGFβR2 interaction is identified as the key molecule in the regulatory functions of tumor-infiltrating B cells in GBM [130]. Furthermore, researchers have attempted to alter the regulatory effect of B cells through stimulating B cells with CD40 agonists, IFN-γ and BAFF to enhance the antigen presentation function of TIBs, thus facilitating antitumor response in GBM [129]. In general, analysis of resection specimens from IDH1/2 wild-type GBM has shown that the association between CD20+Ki67+ B cells and prolonged survival is not significant. Furthermore, GBM contains focal B and T cell aggregates that resemble immature TLS [131]. Although the presence of TLS assembled within brain tumors may serve as supplementary sites for antigen presentation and T cell initiation, the efficiency of tumor immune cycle in the central nervous system is weaker than that in the peripheral nervous system (Figure 4).
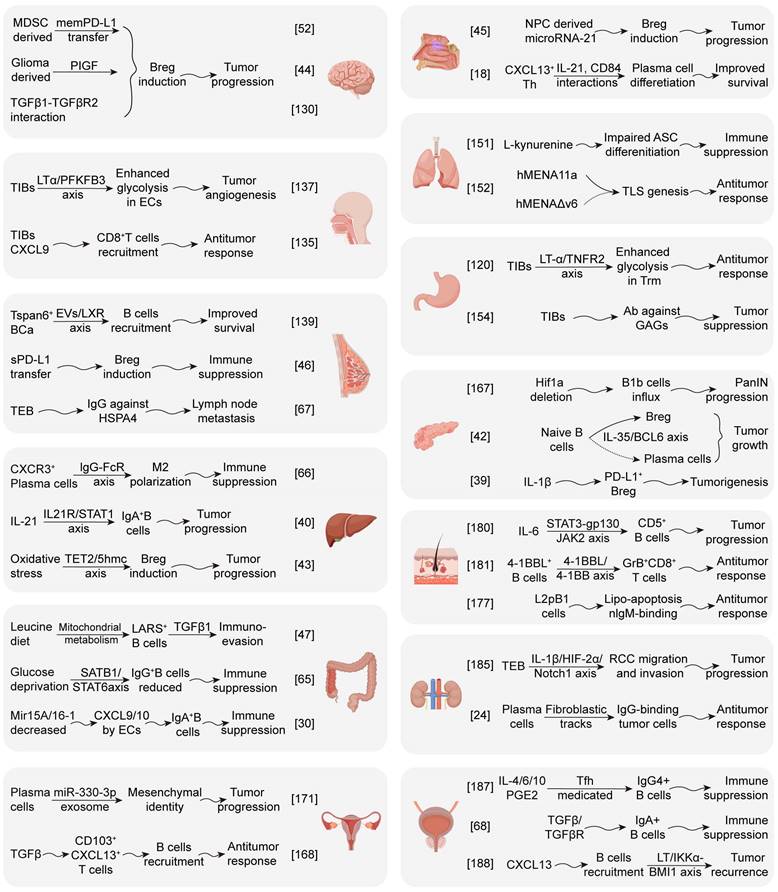
Overview of partial mechanism research on B cells and subsets in the pan-cancer field. The recruitment, differentiation, and effector functions of TIB subsets significantly impact immune response and tumor progression. Solid lines indicate promotion, and dashed lines indicate inhibition (By Figdraw).
Nasopharyngeal carcinoma
Nasopharyngeal carcinoma is an epithelial cancer originating from the mucosal layer of the nasopharynx and is characterized by Epstein-Barr virus (EBV) infection and regional epidemiology. The intense immune infiltration with poor differentiation of NPC is a unique feature that significantly distinguishes NPC from other cancers [132]. Latent EBV infection is typically established in host B lymphocytes and epithelial cells, resulting in an increase in B-cell clonality. Importantly, significant and positive associations are observed between patient survival and the signatures of plasma cells in NPC cohorts [93]. Li et al. demonstrated that TIBs recruited by CXCL13+ Th cells could induce plasma cell differentiation and anti-tumor immunoglobulin production through IL-21 and CD84 interactions in TLS [18]. Notably, elevated serum IgA levels indicate continuous exposure to EBV and serve as high-risk biomarkers of NPC, whereas elevated titers of EBV neutralizing antibodies and anti-gp350 antibodies are considered low-risk biomarkers for NPC. These data suggest that vaccines targeting EBV-gp350 may reduce the risk of NPC. The identification of MEF2B, EBF1, and IL-6R as direct gene targets of EBV nuclear antigen 1 is critical for the survival of B lymphocytes in NPC [133]. Nevertheless, researchers have found that NPC-derived miR-21 induces the formation of IL-10+ B cells, capable of suppressing the activities of CD8+ T cells. Therefore, miR-21 may represent a potential target for the treatment of NPC [45]. In addition, there are studies showing that DNBs participate in regulating immune responses and shaping a suppressive TME in NPC [134]. For further research on plasma cells, Bregs, and DNBs within the TME may reveal a broader role for B cells in NPC.
Squamous cell carcinoma of head and neck
Head and neck squamous cell carcinoma refers to malignant squamous epithelial tumors, that typically arise from exposure to environmental carcinogens or human papillomavirus (HPV) infections. In oropharyngeal squamous cell carcinoma, TIBs may recruit CD8+ T cells through CXCL9 and high-density CD20+ B-cell/CD8+ T-cell aggregation is correlated with good prognosis in patients [135]. In HPV+ HNSCC, researchers have revealed that TIBs contain a high proportion of activated, antigen-presenting, and memory B cell characteristics through flow cytometry. Patients have transcriptional signatures of GC BCs and spatial organization of immune cells consistent with TLS with GCs, both of which correlate with favorable outcome [94,95]. The presence of tumor-specific B cell responses is marked by ASCs in the tumor stroma, which actively synthesize specific IgG antibodies against the HPV proteins E2, E6, and E7 in situ. Studies have shown that LT-α can promote tumor-associated TLS formation, and potentiate recruitment of CD8+ T cells and anti-tumor effects in HNSCC [136].
Although TLS and B cells show positive prognostic effects in HNSCC patients, the functions of regulatory B cell subsets have also been revealed by some researchers. Yang et al. reported that LT-α secreted by TILs promotes proliferation, migration and angiogenesis of endothelial cells by enhancing TNFR/NF-κB/PFKFB3-mediated glycolysis, which may lead to aberrant tumor angiogenesis and progression in HNSCC [137]. In addition, multiple immunosuppressive mechanisms of regulatory B cells have been identified through ADO production in the TME, inhibiting T cell function [138]. The inhibitory effect of ibrutinib on Bruton's tyrosine kinase (BTK) reduces the ADO production by downregulating CD39, significantly increasing B cell infiltration and impede tumor progression in HNSCC.
Breast cancer
Breast cancer, which can be divided into hormone receptor-positive breast cancer, human epidermal growth factor receptor 2-positive breast cancer and triple-negative breast cancer (TNBC), is the most common malignancy among women worldwide. TIBs can sustain a humoral immune response in breast cancer and help generate effective anti-tumor immunity at the tumor situ. It is found that B lymphocytes could be recruited by tumor-derived extracellular vesicles (CCD-EVs) in a liver X receptor-tetraspanin 6 (LXR-Tspan6) dependent manner [139]. A single-cell landscape of breast cancer has shown that intratumoral B cells are primarily naïve or memory cells, rather than ASCs [140]. Compared with peripheral blood, they are more mature, have greater clonality, and greater CSR and SHM characteristics [96]. Using immune receptor sequencing and RNA-seq, researchers found that B cell and T cell responses appear to coevolve with metastatic cancer genomes and reflect tumor mutations and neoantigen structures in breast cancer [141].
Notably, serum-derived soluble PD-L1 can facilitate PD-1+ regulatory B cells induction and IL-10 secretion, thus mediating the immune suppression in invasive breast cancer [46]. Several studies have been conducted on the progression of TDLNs in breast cancer. Tacconi and his colleagues revealed that CD169+ macrophages, the predominant subtype in naïve lymph nodes, exert anti-metastatic effects with the presence of B cells [142]. However, tumor-educated B cells (TEB) accumulate in TDLNs and selectively promote breast cancer lymph node metastasis by producing pathogenic IgG against HSPA4 [67]. In patients receiving neoadjuvant therapy, a positive feedback loop between activated CD24hiCD27+ Bregs and residual tumor cells in the TDLNs promotes multidrug resistance in breast cancer cells [143]. Another study demonstrated that TNFhiIL10+ B cells in TDLNs showed an inverse correlation with the involved lymph nodes, indicating good clinical outcomes in patients with breast cancer [144].
Lung cancer
Lung cancer, including non-small cell lung cancer and small cell lung cancer, is one of the most commonly diagnosed cancers and the leading cause of cancer-related death worldwide. Indeed, naïve-like and plasma-like B cells dominate the TME of lung cancer. A high plasma cell infiltration often signifies a poorer prognosis for LUAD patients [118], whereas it demonstrates greater sensitivity to drug treatments [145]. IHC evaluation of LUAD tissue samples revealed that B cells and plasma cells were observed almost exclusively in the invasive tissue stroma [146]. Similarly, another analysis of lung cancer tissue revealed that intratumoral B cells and plasma cells were mainly localized in the tumor TLS [147]. Naïve-like B cells inhibit the growth of lung cancer cells by secreting molecules that negatively regulate cell growth, whereas plasma-like B cells inhibit cancer cell growth in the early stages of NSCLC but promote cell growth in the advanced stages of NSCLC [148].
In addition, some studies have shown that high frequencies of CD24hiIL-10+ Bregs with plasmacytoid gene signatures are found in lung cancer patients [149]. CD27-lgD-double-negative B cells in NSCLC have also been identified and shown a negative correlation with the presence of mature B cell populations [150]. In flamed NSCLC, IDO1 is upregulated in cancer cells and TLS, increasing tryptophan degradation and metabolite kynurenine production, thus mediating an immunosuppressive effect [151]. Moreover, a study by Modugno et al. indicated that NSCLC tumoral hMENA11a upregulated LTβR, decreased fibronectin, and favored CXCL13 production and TLS formation by Trms. In contrast, hMENAΔv6 in cancer-associated fibroblasts inhibited LTβR-associated TLS formation via NF-kB pathway [152].
Gastric cancer
Gastric cancer is the fifth most common cancer and the third leading cause of cancer death worldwide. High levels of CD20+ B cell infiltration are significantly associated with improvements in overall survival and disease-free survival of GC patients [97]. Most infiltrating B cells in gastric cancer around the tumor margin in the form of TLS are sensitized by antigens and serve as APCs in TLS, participating in the induction of cytotoxic T cells [153]. Researchers have revealed that sulfated glycosaminoglycans (GAGs) are the major functional B cell antigens in gastric tumors. Utilizing natural anti-sulfated GAG antibodies inhibits the growth of various human malignancies [154]. Another study indicated that CD103+ T cells in the tumor epithelium were located around TLS and CD103+CD8+ Trm cells in tumor were associated with the TLS, leading to the enhancement of anti-tumor immunity in gastric cancer [155]. TLS associated B cells could promote glycolysis of CD103+CD8+Trm cells through the LT-α/TNFR2 axis, enhancing the secretion of CXCL13, granzyme B and antitumor immunity in gastric cancer tissues [120].
In addition, Hu et al. found that IL-10-expressing CD27+CD10- B cells aggregate in the intratumoral environment of gastric cancer, which may significantly reduce the production of IFN-γ, TNF, and IL-17 and inhibit antitumor response [156]. Similarly, CD19+CD24hiCD27+Bregs could inhibit IFN-γ production of CD4+T cells and promote immune escape in gastric cancer [157]. Furthermore, it is reported that Bregs in the cancerous mucosa and PBMC increase with cancer stage and promote gastric carcinogenesis by inducing the production of inflammatory mediators IL-10 [34]. With the levels of Tfh, Breg, and CXCL13 increased, related lymph node metastasis and poor prognosis have been observed in gastric cancer patients.
Liver cancer
Liver cancer remains a global health challenge, with hepatocellular carcinoma as the most common form. In the past decade, several studies have been conducted on functional B cells and Breg subsets of TIBs in HCC. The close proximity of TILs and B cells is associated with heightened intratumor immunity and contribute to an improved prognosis for patients with HCC [75]. Importantly, the refined immunosubtype, derived from the spatial dynamics of T and B cell responses, can effectively predict the clinical trajectory following surgical resection of resectable HCC and Atezo/Bev therapy for advanced HCC [158]. In addition, the immunogenic subtype of intrahepatic cholangiocarcinoma is characterized by inflammatory infiltration and is associated with prolonged patient survival, featuring a variety of immune cells, including effector and memory T cells, B cells, and macrophages [159]. Intra-tumoral TLS are linked to a reduced risk of early relapse following surgical resection and effective in situ anti-tumor immunity [112]. These results suggest that local immunity is activated in the early stage of HCC.
Researchers uncovered that CXCR3+ B cells account for approximately 45% of TIBs and related to early recurrence of human HCC [16]. The crosstalk elucidated between macrophages and CXCR3+IgG+ plasma could promote inhibitory cytokines secretion of macrophages and diminish the effect of CD8+ T cells in the murine models of HCC [66]. Furthermore, it has been reported that the IL-21R-STAT1 axis is activated in MASH-driven HCC, leading to the induction of immunosuppressive IgA+ plasma cells and tumor progression [40]. Several intervention strategies involving Breg have been reported. Inhibiting the catalytic activity of ten-eleven translocation-2 (TET2) enzymes in B cells can enhance antitumor immunity and improve the efficacy of PD-1 therapy in HCC [43]. In particular, it has been suggested that the ablation of B cells by CD20 antibodies can limit hepatic fibrosis-driven tumorigenesis in HCC mouse models. Besides, high infiltration of FcγRIIlow/- B cells activated by semimature DCs is linked to cancer progression and poor prognosis, whereas the interaction between tumor-infiltrating T cells and B cells has also been identified and improved outcomes in HCC patients [75,84].
Colorectal cancer
In recent years, the incidence and mortality rates of CRC have increased globally. Mature TLS and plasma cells have been reported in CRC or adjacent tissue, which are associated with improved clinical outcomes in patients. In colorectal cancer liver metastasis, CCL19+ fibroblasts have been found to facilitate lymphocyte recruitment to TLS, thereby enhancing the antitumor immunity of IgG+ plasma cells [98]. CD86+ antigen-presenting B cells were identified in the GC of TLS and isolated from TDLNs, inducing responses from autologous T cells in vitro [160]. Human IL-36γ was associated with CD4+ central memory T cell infiltration, increased B cell density in TLS, and markers of fibrosis, supporting IL-36γ as the physiological immune responses through maintaining inflammation in CRC [161]. It is reported that declined micro15A/16-1 levels in tumors increased I-kappaB kinase-mediated NF-κB/STAT1 activation, which lead to the production of chemokines CXCL9 and CXCL10 of epithelial cells and promote the progression of CRC through chemotaxis of IgA+IL-10+ plasma cells [30]. Wang et al. demonstrated that LARS2+ B cell subset shows a regulatory phenotype and is located outside the TLS, while leucine starvation could inhibit LARS2 B cell-mediated immune evasion in CRC [47]. Moreover, researchers have revealed that the deceleration of glucose metabolism in TIBs suppresses IgG+ plasma cell differentiation through the SATB1 pathway, whereas intervention in glucose metabolism may enhance the anti-tumor effect of B cells [65].
Pancreatic cancer
Pancreatic cancer has the worst prognosis among all common solid malignancies, with a 5-year overall survival rate of approximately 10%. Exocrine pancreatic cancer accounts for approximately 95% of all pancreatic cancer cases, with the most common type being ductal adenocarcinoma. PDAC exhibits a highly suppressive TME and significant B cell infiltration, characterized by extensive inflammatory fibrosis, desmoplastic stromal reaction, and hypo-vascularity [162]. Further analysis of PDAC tissue shows that groups with mature TLS support plasma cell differentiation as well as the formation of tumor-reactive T cells [163]. In the peripheral blood of PDAC patients, circulating plasmablasts proliferate and infiltrate pancreatic lesions, stimulating collagen production in fibroblasts and participating in extracellular matrix remodeling [28]. Autoantibodies targeting filamentous actin and nuclear protein RuvB-like AAA ATPase2 antigens have been detected in the TME and peripheral blood, supporting the initiation of humoral immunity in the chronic inflammatory microenvironment of PDAC [164]. Capello et al. showed that tumor antigens released by exosomes from PDAC could trigger the production of autoreactive antibodies and serve as decoys for complement-mediated cytotoxicity [165].
In addition, various phenotypes of IL-35 producing Bregs including CD1dhiCD5+ B cells and CD38+ B cells have been reported in PDAC [37,54,166]. Studies have shown that IL-1β of TME promotes tumor progression through increasing CXCL13 expression and infiltration of immune-suppressive IL-35+ B cells [39]. Dysregulated transcriptional programs resulting from IL-35-induced BCL6 upregulation in B cells, impair the differentiation of naïve B cells into antitumor plasma cells through the stimulation of STAT3-PAX5 complex in PDAC [42]. For B-cell-mediated IL-35 induction, BCR stimulation of BTK signaling is necessary and inhibition of the key downstream molecule PDK2 enhances effector T cell function [54,55]. Researchers have uncovered that IL-35 production by circulating B cells in PDAC indicates CD8+ T cell exclusion and immunotherapy resistance via IL-35/gp130/STAT3 [35]. Furthermore, hypoxia-inducible factor 1α (HIF1α) has been identified as a protective factor in the preinvasive phase of PDAC, since its absence causes pro-tumor B1b influx and tumor progression [167].
Ovarian cancer
Ovarian cancer is the second most common cause of gynecological cancer-related death among women worldwide. Analysis of tumors and stromal cells reveals that intratumoral plasma cells are associated with a better prognosis in patients with HGSOC [99]. Ovarian clear-cell carcinomas with an inflammatory stroma constitute a unique clinicopathological subgroup, and these tumor cells induce inflammation and stimulate plasma cell differentiation in a paracrine manner. TGFβR signaling licensed CD8+ T cells upregulate CD103 and secrete CXCL13 with TCR stimulation, involved in mediating B cell recruitment and TLS formation in ovarian cancer [168]. Studies have pointed out that IgA-binding tumor antigens can mediate the transcytosis and cytosis, promoting cytotoxic T lymphocyte (CTL) killing of tumor cells and inhibiting tumor growth in ovarian cancer through myeloid cell-dependent mechanisms, neutralizing secreted factors, or poly-Ig receptors on cancer cells [60]. Intratumoral ASCs also produce tumor-reactive IgG targeting MMP14 in HGSOC, specifically encapsulating tumor cells with high MMP14 expression [169]. In omental metastases of HGSOC, B cells primarily infiltrate lymphoid structures in the stroma. Tumor-specific IgG is produced and mediates a strong B cell memory response against a limited antigen repertoire, supporting antitumor response [170]. T cells and B cells colocalized in lymphoid aggregates, ranging from small diffuse clusters to well-organized TLS, suggesting that synergistic interactions could generate and enhance the effective antitumor immunity in HGSOC [74].
However, Yang et al. found that the switch of HGSOC-mesenchymal phenotype can be triggered by the transportation of plasma cell-derived exosomes containing miR-330-3p, which increases the expression of junctional adhesion molecule B in an atypical manner and is associated with tumor development [171]. In addition, Bregs has been reported to preferentially enrich ascites in ovarian cancer and impair anti-tumor immunity [172]. In the ovarian cancer murine model, peritoneal ascite-B1 cells, spleen B1 cells and marginal zone B cell subsets were selectively lost due to the influx of MDSCs [173]. In fact, mature TLS are formed only in 16% of HGSOC with a relatively high TMB [119]. It is well-supported that the anti-tumor immune effects of humoral immunity and tumor-associated TLS in ovarian cancer have not been fully elucidated.
Melanoma
Cutaneous melanoma is a malignant tumor that arises from melanocytes in the skin. Current evidence supports that B cells enhance tumor immunity against B16F10 melanoma by promoting T cell infiltration into tumors and cytokine production in the TME, associated with better prognosis in melanoma [174]. When exposed to melanoma secretomes, B cells express inflammatory factors, T cell-recruiting chemokines, differentiating into plasmablast-like cells to sustain inflammatory state [175]. The co-occurrence of tumor-associated CD8+ T cells and CD20+ B cells is associated with improved survival, independently of other clinical variables [176]. In addition, L2pB1 cells, as a subpopulation of B1b cells, have been reported to inhibit melanoma growth by inducing lipo-apoptosis of tumor cells [177]. Immature and follicular B2 cells exert protective effects against melanoma progression by promoting the generation of effector memory T cells [178]. As the macrophage barrier is disrupted with the application of therapeutic agents, the contact of tumor-derived vesicles with B cells can enhance tumor-related humoral immunity in TDLNs [82].
Extensive findings underscore the diverse roles of B cells in the peripheral blood, TME, and TDLNs of melanoma. In addition, IL-10+ B1a cells selectively accumulate in melanoma and promote tumor growth by inhibiting tumor-infiltrating CD8+ T cells [38]. In TDLNs of B16F10 melanoma, researchers have identified a subset of TIM-1+ B cells, while selective deletion of TIM-1 enhances the IFN-γ response and anti-tumor immunity [179]. Animal experiments have shown that IL-6 binds to CD5+ B cells and can activate the transcription factor STAT3 through gp130 and its downstream kinase JAK2, thereby promoting the progression of melanoma [180]. Lee-Chang et al. proposed that 4-1BBL+ B cells could increase antitumor responses in aged mice by presenting endogenous antigens and inducing GrB+CD8+ T cells via the 4-1BBL/4-1BB axis [181].
Renal clear cell carcinoma
Renal cell carcinoma (RCC) is the most common form of kidney cancer and is typically divided into clear cell carcinoma (ccRCC) and non-clear cell renal cell carcinoma. B cell repertoire analysis revealed clonal diversification, selection, and expansion of B cells, along with the prolonged PFS and response to ICI in TLS+ ccRCC. Further studies have shown that IgG- and IgA-producing B cells spread into the tumor bed along fibroblast tracks in TLS [24]. Analysis of genomic alterations in TLS shows that the PI3K-mTOR pathway and the proportion of PD-1+ cells are generally upregulated in TLS+ tumors of ccRCC [182]. Xu et al. showed that most TLS in ccRCC are located in the distal areas of the tumor and are associated with immature and immunosuppressive characteristics, whereas tumor-adjacent TLS correlate with favorable clinical outcomes [183]. Similarly, another study also supported that the high density of intratumoral TLS is an independent risk factor for a favorable prognosis in ccRCC [184]. Besides, it is reported that B cells recruited by tumor cells can activate IL-1β/HIF-2α signaling and induce the downstream Notch1 signaling pathway, significantly increasing the migration and invasion of RCC cells [185].
Prostate cancer
Prostate cancer progresses from prostatic intraepithelial neoplasia to locally invasive adenocarcinoma, and eventually to castration-resistant metastatic cancer. Prostate tumors in Black men of African descent exhibit enhanced plasma cell infiltration, elevated markers of NK cell activity, and increased IgG expression, which are associated with better outcomes [100]. Studies have shown that B cell infiltration is higher in intratumoral areas than the peritumoral areas of prostate cancer tissues during prostatectomy [186]. The proportion of Tfh2 and IgG4+ B cells in patients with prostate cancer is increased compared with healthy peripheral blood. Further studies have shown that cytokines, such as IL-4, IL-6, IL-10 and PGE2, can promote the antibody class switching of B cells to IgG4, partly through the induction of Tfh2 cells, which is related to the immunosuppressive microenvironment [187]. In addition, it has been found that the CXCL13 recruits B cells into prostate cancer tumors and activate the IκB kinase α-BMI1 module in cancer stem cells through the production of LT, thus promoting the progression of castration-resistant prostate cancer [188]. Shalapour et al. identified the key TGFβR signaling immunosuppressive B cells in prostate cancer as plasma cells expressing IgA, IL-10, and PD-L1, which are involved in tumor immune tolerance [68].
Prospects of B cells and TLS in tumor immunotherapy
Immunotherapy has become a new pillar of cancer treatment through blocking receptors and ligands that weaken T cell activation pathways, reversing the state of T cell exhaustion in the TME. Drug resistance and immune-related adverse events (IRAEs) limit the further application of ICB. There is evidence that B cells and antibodies can serve as predictors of the IRAEs during ICB therapy [189]. Given the observed link between TLS and clinical outcomes in various cancer, targeting B cells and manipulating TLS may present an appealing therapeutic approach.
B cells and antibodies-based tumor treatment strategies
Preclinical research and clinical applications focuses on B cell treatment strategies including depletion of Breg, anti-tumor B cell activation, antibody production, cytokine therapy and B cell vaccines. B cell depletion can be used to treat autoimmune diseases, infections and malignant B cell diseases. Drugs for Breg depletion through CD20 antibodies pose risks and controversies in cancer treatment, given the association of B cells with a better prognosis in most tumors [190]. Nano-capture technology of CXCL13 could reduce tBreg differentiation and normalize epithelial-mesenchymal transition in the TME, thereby inhibiting tumor growth and prolonging the PFS in pancreatic cancer, BRAF mutant melanoma, and TNBC murine models [191]. Intratumoral delivery of CCL21 nanoparticles has been associated with the inhibition of lung cancer growth, reflecting the complexity of chemokine-based therapies [192]. In addition, it is reported that nano-delivery therapy of PI3K-γ inhibitors enhance the anti-tumor immunity by hampering cellular activation and migration to reduce suppressive myeloid and plasma cell infiltration in tumors [193]. Inhibiting the BTK of B cell can restore T cell-dependent anti-tumor responses and enhance sensitivity to chemotherapy treatments [194]. The BTK inhibitor Ibrutinib impedes PDAC progression and improves response to chemotherapy, providing a new treatment for pancreatic cancer.
Engineered antibodies have significantly expanded the strategic spectrum of targeted therapy and immunotherapy, providing promising perspectives for cancer treatment [195]. Monoclonal antibodies are developed for ICB therapies, anti-angiogenic drugs, antibodies targeting tumor antigens, and antibody-drug conjugates incorporating drugs, radioisotopes and immunotoxins, while bispecific antibodies are also employed in cancer treatment through targeting T cells plus tumor epitopes or double tumor antigen epitopes (Figure 5). There are various studies reported B cell activation components (IL-2, IL-6, IL-15, IL-21, BAFF, APRIL, rhIL-12, fucose and CXCL13-coupled CpG-ODN) and Breg suppressive agents (lipoxin A4, MK886, total glucosides of paeony and resveratrol) to regulate the proportion of functional B cells in TME [49]. Under the concept of therapeutic cancer vaccines, activation of 4-1BBL+ B cell vaccines with CD40 agonism and IFN-γ elicits potent immunity effects in GBM murine models by enhancing antigen-presenting and antibody-secretion capabilities through in vitro stimulation and reinfusion [129]. These studies highlight the potential applications of B cells and antibodies for cancer treatment, emphasizing the need to enhance anti-tumor B cells and antibodies while concurrently inhibiting the Breg phenotype.
B cell and antibody-based therapeutic strategies. Current B cell therapy in cancer is primarily utilized in preclinical stages within in vitro and animal models, whereas engineered antibodies have been widely adopted in clinical applications due to their antigen-specificity. TGP: total glucosides of paeony; LXA4: lipoxin A4.
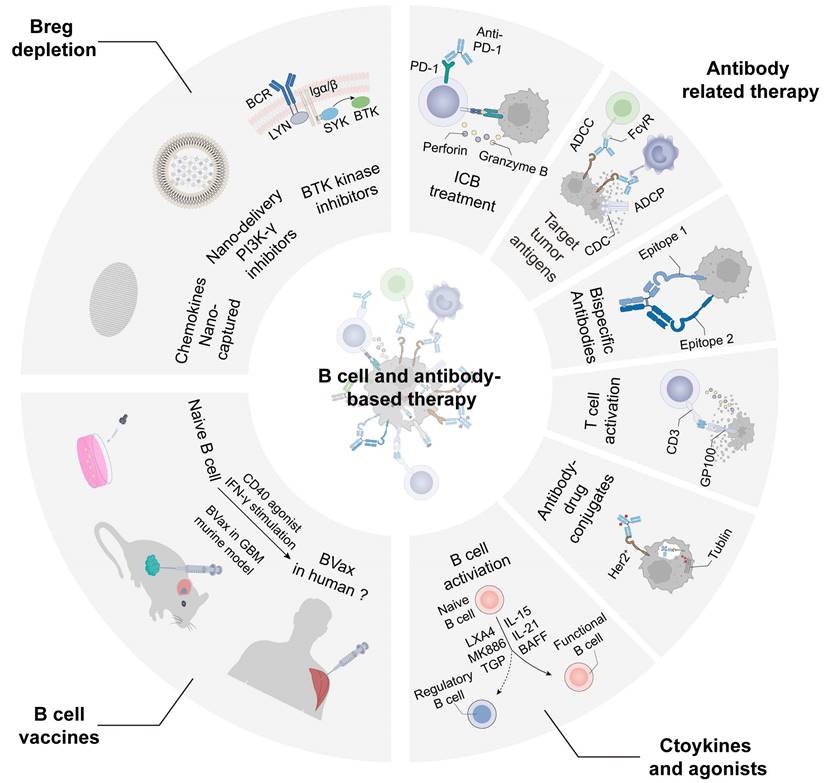
TLS-based tumor treatment strategies
Based on the positive impact of TLS in the prognosis of tumor patients, devising methods to induce TLS formation has become an effective strategy. Several studies have shown that enhancing tumor-related TLS can broaden the scope of current cancer treatment in various systemic and intratumoral manners. Among systemic therapies, approaches such as radiotherapy [196] and chemotherapy [197] in mice and human have shown the potential to induce TLS, which are essential for maintaining the long-term responses to immunotherapy [198]. While enhancing tumor immunity by increasing tumor-associated TLS is effective through systemic therapies, current intratumoral interventions need to specifically target and manipulate the TME. In murine models, it has been reported that increased levels of chemokines [199], cytokines (LT, TNFα, LIGHT-VTP, CXCL13/CCL21, IL-15, TGF-β, etc.) [200] and HEV have been reported to be involved in TLS genesis [110]. The use of agonists to activate pathways that normalize tumor vasculature and enhance immune infiltration. Many studies have referred to the role of agonist agents [201]. Oncolytic adenoviruses [202] and humanized cancer vaccines [203] serve as delivery systems for therapeutic agents to facilitate the genesis and function of TLS in tumors.
In addition, biomaterial materials including 3D scaffolds [204], engineered DCs [205], collagen matrices [206], hydrogels [207], nanomaterials [208] and organ-on-a-chip/organoids [209] have been developed to mimic and enhance immune structures within tumors. These approaches aim to establish a supportive environment for the formation of TLS, thereby increasing the capacity of the immune system to combat cancer. Collectively, these strategies exemplify multifaceted approaches to harnessing TLS in the pursuit of more effective cancer therapeutics (Table 4). Since TLS have the potential to induce the excessive activation of anti-tumor immunity in situ, it is necessary to consider the methods and risks that pan-specific manipulation of TLS may exacerbate IRAEs.
TLS induction strategies in mouse models and cancer patients
| Categories | Treatment | Subject Type | Description | Refs |
|---|---|---|---|---|
| Systemic therapies | Radiotherapy | Mice | TLS with medullary breast cancer is described in a patient case report, and similar to TLS observed in the genetic mouse model. | [196] |
| Chemotherapy | Patient | Chemotherapy induces C3 cleavage and CR2 activation, resulting in CR2+ B cells expressing ICOS-L, which accumulate in TLS. | [197] | |
| Immunotherapy | Patient | TLS may be involved in initiating and maintaining long-term responses to ICB. | [101, 125, 147] | |
| Targeted therapy: Light-VTP | Mice | Targeting LIGHT to tumor vessels via VTP leads to the formation of TLS and prolongs survival in mice when combined with ICB treatment. | [198] | |
| Intratumor therapies | Chemokines: CXCL13/CXCR5 CCL21/CCR7 | Mice | Under the control of the rat insulin, expression of LTα1β2 promoter induces HEV formation and secretion of CCL19/21, and CXCL13 in pancreatic tissue, leading to the development of TLS. | [199] |
| Cytokines: Lymphotoxin-α Lymphotoxin-β | Mice | CD8+ T cells, together with TIBs that secrete lymphotoxin-α1β2 (LTα1β2), act as LTi cells and coordinate the formation and expansion of TLS. | [200] | |
| Agonistic agents: STING agonists TLR4 agonists CD40 agonists | Mice Mice Mice | Low doses of the STING agonist injection have been shown to promote tumor vascular normalization, increase immune infiltration, and induce the formation of immature TLS. | [201] | |
| Oncolytic virus | Tumor-selective oncolytic virus vector | Mice | The identified 4-E03 antibody via oncolytic vaccinia vector exhibits stronger Treg depletion against Treg cells expressing CTLA-4, potentially enhancing the functionality of TLS. | [202] |
| Humanized cancer vaccine | (GM-CSF)-secreting PDAC cell vaccine (GVAX) | Patient | Compared with tumors from group of unvaccinated patients, patients vaccinated intradermally with irradiated GCAX induce an increase in TLS formation of tumor. | [203] |
| Biomaterials | 3D TLS scaffold | Mice | The 3D scaffold with a porous structure can recruit numerous immune cells to endow it with functions similar to real lymphatic organs, thus forming an artificial tertiary lymphatic structure. | [204] |
| Engineered DCs | Mice | T-bet and IL-36γ synergistically enhance each other's expression in DCs, enabling them to promote TLS formation in the “immune normalization” therapeutic TME. | [205] | |
| Collagen matrices | Mice | Transplantable and functional TLS are constructed by applying soluble factors, encapsulated within a slow-release gel. | [206] | |
| Hydrogels | Patient | By combining Zn2+ with 4,5-imidazole dicarboxylic acid and incorporating chitosan and CpG, a STING-activating hydrogel (ZCCG) is meticulously prepared to initiate and activate cGAS-STING and TLR9 pathway-mediated immunotherapy. | [207] | |
| Nanomaterials | Mice | A nano-vaccine, composed of EBNA1 and a bi-adjuvant combination of Mn2+ and CpG formulated with tannic acid, enhances the expression of downstream chemokines CCL19/21, and CXCL13 by activating the LT-α and LT-β pathways to promote the formation of TLS. | [208] | |
| Organ-on-a-Chip/organoids | Vitro | Primary B and T cells isolated from human blood are cultured in a 3D extracellular matrix gel and are able to independently assemble into ectopic lymphoid follicles. | [209] |
VTP: vascular targeting peptide; T-bet: T-box expressed in T cells; CPG: cytosine phospho-guanine; EBNA1: Epstein-Barr virus nuclear antigen 1.
Conclusions and perspectives
Particularly, surgical resection, alleviating the immunosuppressive TME, helps achieve an equal fight between immune cells and residual tumor cells. Postoperative immunotherapy and targeted therapy can support the individual immune system and effectively eliminate residual tumor cells and micrometastases. However, postoperative drug resistance, along with tumor recurrence and metastasis, continues to pose significant challenges in the treatment of solid tumors. It should be pointed out that surgical treatment removes tumor tissue but also terminates the intratumoral TLS effect. Although CXCR6+ tissue-resident memory T and B cell clones clustering with CXCL16+ DCs allows the survival of transitory effector state TCF1-CX3CR1+ CTLs, they may be not adequate for response to tumor recurrence and still require additional circulating recruitment [15]. In essence, tumor recurrence and metastasis can be interpreted as the insufficiency of local priming and the hysteresis of circulating immunity. The time difference between the individual response to tumor specific antigens and the proliferation of residual tumor cells determines the possibility and risk of recurrence after surgery, which is analogous to the tortoise and the hare race. It is particularly important to establish effective local immunity and immunological memory to eradicate remaining tumor cells through biocompatible engineered materials after surgery. Therefore, utilizing engineered antibodies, therapeutic tumor vaccines and establishing biocompatible mature TLS may potentially make up for this time difference and provide a higher response likelihood for tumor antigens that lymphocytes encounter (Figure 6). Future treatments need to explore effective strategies for cancer treatment from multiple dimensions including intensity and speed to attain more therapeutic benefits.
Postoperative models of individual immune response and tumor proliferation. (A) Surgical resection removes tumor tissue while also terminating the local immunity of intratumoral TLS. (B) Postoperative immunity targeting residual tumor cells requires the reactivation of circulating activation pathways. The hysteresis in circulating immunity and the re-establishment of local immunity provide the opportunity for rapid tumor growth. The speed difference of the individual immune response and tumor growth determine the status of tumor development.
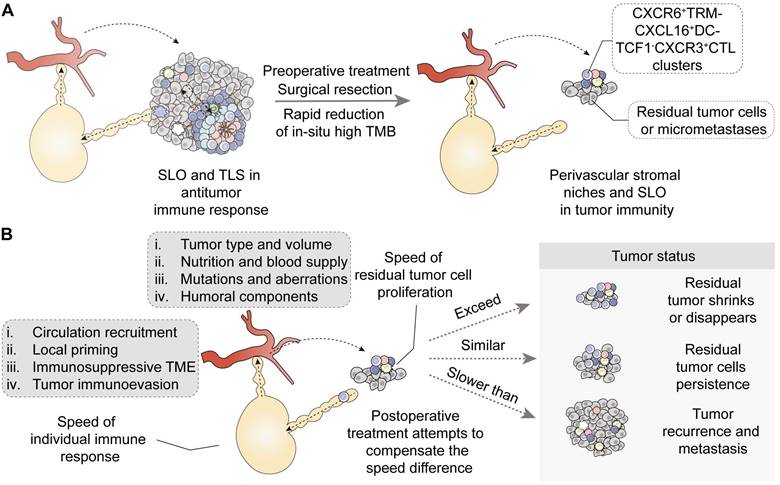
Abbreviations
AHR: aryl hydrocarbon receptor
AtM B cell: atypical memory B cell
ADCC: antibody-dependent cellular cytotoxicity
ADO: adenosine
ADCP: antibody-dependent cellular phagocytosis
APC: antigen-presenting cell
ASC: antibody secreting cell
Breg: regulatory B cell
BAFF: B-cell activating factor
Breg: regulatory B cell
BTK: Bruton's tyrosine kinase
CCL21: CC-chemokine ligand 21
ccRCC: clear cell renal cell carcinoma
CDC: complement-dependent cytotoxicity
CCD-EV: cancer cell-derived extracellular vesicle
CLP: common lymphoid progenitor
CPG: cytosine phospho-guanine
CRC: colorectal cancer
CSR: class switch recombination
CTL: cytotoxic T lymphocyte
CXCL13: CXC-chemokine ligand 13
DFS: disease-free survival
DNB: double negative B cell
DSS: disease-specific survival
EBNA1: Epstein-Barr virus nuclear antigen 1
EBV: Epstein-Barr virus
EBF1: early B-cell factor 1
EF: extrafollicular
EF BC: extrafollicular B cell
ERV: endogenous retrovirus
FFS: failure-free survival
FDC: follicular dendritic cell
FRC: fibroblast reticular cell
FOXO1: Forkhead Box O1
GABA: γ-aminobutyric acid
GAG: glycosaminoglycan
GBM: glioblastoma
GC BC: germinal center B cell
GrB: granzyme B
H&E staining: hematoxylin and eosin staining
HA: hyaluronic acid
HCC: hepatocellular carcinoma
HGSOC: high-grade serous ovarian cancer
HEV: high endothelial vein
HIF1a: hypoxia-inducible factor 1α
HMGB1: high mobility group box 1
Hhep: helicobacter hepaticus
HNSCC: head and neck squamous cell carcinoma
HSC: hematopoietic stem cell
HSPA4: heat shock protein 4
ICB: immune checkpoint blockade
IDO: indoleamine-2,3-dioxygenase
IF: immunofluorescence
IGKC: immunoglobulin kappa C
ILC3: group 3 innate lymphoid cell
IL-10: interleukin 10
IRAE: immune-related adverse event
IRF4/8: interferon regulatory factor 4/8
LARS2 B: leucine-tRNA-synthetase-2-expressing B cell
L-kyn: L-kynurenine
LMA: lympho-myeloid aggregate
LN: lymph Node
LT: lymphotoxin
LTB4: leukotrienes B4
LTi: lymphoid tissue inducing cell
LTo: lymphoid tissue organizer cell
LUAD: lung adenocarcinoma
LXA4: lipoxin A4
LXR-Tspan6: liver X receptor-tetraspanin 6
MASH: metabolic dysfunction-associated steatohepatitis
MDSC: myeloid-derived suppressive cell
mIHC: multiplex immunohistochemistry
MZ BC: marginal zone B cell
NK cell: natural killer cell
NPC: nasopharyngeal carcinoma
NSCLC: non-small cell lung cancer
OS: overall survival
PAX5: Paired Box 5
PBMC: peripheral blood mononuclear cell
PDAC: pancreatic ductal adenocarcinoma
PD-L1: programmed death ligand 1
PFS: progression-free survival
PlGF: placental growth factor
PNAd: peripheral node addressin
PPARα: peroxisome proliferator-activated receptor α
Pre-B cell: precursor B cell
Pro-B cell: progenitor B cell
RCC: renal cell carcinoma
RFS: recurrence-free survival
ROS: reactive oxygen species
RNA-seq: RNA sequencing
SHM: somatic hypermutation
SLO: secondary lymphoid organ
ST: spatial transcriptomics
TAA: tumor-associated antigen
Tbet: T-box expressed in T cell
tBreg: tumor-induced Breg
TDLN: tumor-draining lymph node
TEB: tumor-educated B cell
TET2: ten-eleven translocation-2
TIM: T-cell immunoglobulin and mucin-domain
Tfh: T follicular helper cells
Tfr: follicular regulatory T cell
TIB: tumor-infiltrating B cell
TIL: tumor-infiltrating T lymphocyte
TLS: tertiary lymphoid structures
TLR: toll-like receptor
TMB: tumor mutation burden
TME: tumor microenvironment
TNBC: triple-negative breast cancer
TNF: tumor necrosis factor
Tph: T peripheral helper cell
Treg: regulatory T cell
TRAIL: TNF-related apoptosis-inducing ligand
Trm: tissue-resident memory T cell
TSA: tumor-specific antigen
VEGFC: vascular endothelial growth factor C
VTP: vascular targeting peptide
12-CS: 12-chemokine signature
5-hmc: 5-hydroxymethylcytosine
5-LO: 5-lipoxygenase
Acknowledgements
This project was supported by the National Natural Science Foundation and Young Scientists Fund of China (Grant No. 82373052 and No.82303185).
Author contributions
Xing Wu contributed to the acquisition of literature, drafting of the original manuscript and drawing the figures. Xiaoping Chen, Bixiang Zhang and Binhao Zhang were responsible for the conceptualization. Junnan Liang prepared the draft, and Qibo Huang substantially revised it. All authors reviewed and approved the final manuscript.
Data availability
No datasets were generated or analyzed during the current study.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Gao X, Shen Q, Roco JA, Dalton B, Frith K, Munier CML. et al. Zeb2 drives the formation of CD11c+ atypical B cells to sustain germinal centers that control persistent infection. Sci Immunol. 2024;9:eadj4748
2. Downs-Canner SM, Meier J, Vincent BG, Serody JS. B Cell function in the tumor microenvironment. Annu Rev Immunol. 2022;40:169-93
3. Laumont CM, Banville AC, Gilardi M, Hollern DP, Nelson BH. Tumour-infiltrating B cells: immunological mechanisms, clinical impact and therapeutic opportunities. Nat Rev Cancer. 2022;22:414-30
4. Grasset EK, Chorny A, Casas-Recasens S, Gutzeit C, Bongers G, Thomsen I. et al. Gut T cell-independent IgA responses to commensal bacteria require engagement of the TACI receptor on B cells. Sci Immunol. 2020;5:eaat7117
5. Schumacher TN, Thommen DS. Tertiary lymphoid structures in cancer. Science. 2022;375:eabf9419
6. Le Rochais M, Hémon P, Ben-Guigui D, Garaud S, Le Dantec C, Pers J-O. et al. Deciphering the maturation of tertiary lymphoid structures in cancer and inflammatory diseases of the digestive tract using imaging mass cytometry. Front Immunol. 2023;14:1147480
7. Jones GW, Hill DG, Jones SA. Understanding immune cells in tertiary lymphoid organ development: it is all starting to come together. Front Immunol. 2016;7:401
8. Seleznik GM, Reding T, Romrig F, Saito Y, Mildner A, Segerer S. et al. Lymphotoxin β receptor signaling promotes development of autoimmune pancreatitis. Gastroenterology. 2012;143:1361-74
9. Fleige H, Ravens S, Moschovakis GL, Bölter J, Willenzon S, Sutter G. et al. IL-17-induced CXCL12 recruits B cells and induces follicle formation in BALT in the absence of differentiated FDCs. J Exp Med. 2014;211:643-51
10. Teillaud J-L, Houel A, Panouillot M, Riffard C, Dieu-Nosjean M-C. Tertiary lymphoid structures in anticancer immunity. Nat Rev Cancer. 2024;24:629-46
11. Overacre-Delgoffe AE, Bumgarner HJ, Cillo AR, Burr AHP, Tometich JT, Bhattacharjee A. et al. Microbiota-specific T follicular helper cells drive tertiary lymphoid structures and anti-tumor immunity against colorectal cancer. Immunity. 2021;54:2812-24
12. Playoust E, Remark R, Vivier E, Milpied P. Germinal center-dependent and -independent immune responses of tumor-infiltrating B cells in human cancers. Cell Mol Immunol. 2023;20:1040-50
13. Sarti Kinker G, da Silva Medina T. Tertiary lymphoid structures as hubs of antitumour immunity. Nat Rev Cancer. 2023;23:803
14. Mellman I, Chen DS, Powles T, Turley SJ. The cancer-immunity cycle: Indication, genotype, and immunotype. Immunity. 2023;56:2188-205
15. Mempel TR, Lill JK, Altenburger LM. How chemokines organize the tumour microenvironment. Nat Rev Cancer. 2024;24:28-50
16. Liu R-X, Wei Y, Zeng Q-H, Chan K-W, Xiao X, Zhao X-Y. et al. Chemokine (C-X-C motif) receptor 3-positive B cells link interleukin-17 inflammation to protumorigenic macrophage polarization in human hepatocellular carcinoma. Hepatology. 2015;62:1779-90
17. J Gunderson A, Rajamanickam V, Bui C, Bernard B, Pucilowska J, Ballesteros-Merino C. et al. Germinal center reactions in tertiary lymphoid structures associate with neoantigen burden, humoral immunity and long-term survivorship in pancreatic cancer. Oncoimmunology. 2021;10:1900635
18. Li J-P, Wu C-Y, Chen M-Y, Liu S-X, Yan S-M, Kang Y-F. et al. PD-1+CXCR5-CD4+ Th-CXCL13 cell subset drives B cells into tertiary lymphoid structures of nasopharyngeal carcinoma. J Immunother Cancer. 2021;9:e002101
19. Grootveld AK, Kyaw W, Panova V, Lau AWY, Ashwin E, Seuzaret G. et al. Apoptotic cell fragments locally activate tingible body macrophages in the germinal center. Cell. 2023;186:1144-61
20. Zhu W, Germain C, Liu Z, Sebastian Y, Devi P, Knockaert S. et al. A high density of tertiary lymphoid structure B cells in lung tumors is associated with increased CD4+ T cell receptor repertoire clonality. Oncoimmunology. 2015;4:e1051922
21. Fridman WH, Meylan M, Pupier G, Calvez A, Hernandez I, Sautès-Fridman C. Tertiary lymphoid structures and B cells: An intratumoral immunity cycle. Immunity. 2023;56:2254-69
22. Ma J, Wu Y, Ma L, Yang X, Zhang T, Song G. et al. A blueprint for tumor-infiltrating B cells across human cancers. Science. 2024;384:eadj4857
23. Cancro MP. Age-Associated B Cells. Annu Rev Immunol. 2020;38:315-40
24. Meylan M, Petitprez F, Becht E, Bougoüin A, Pupier G, Calvez A. et al. Tertiary lymphoid structures generate and propagate anti-tumor antibody-producing plasma cells in renal cell cancer. Immunity. 2022;55:527-41
25. Zhang L, Strange M, Elishaev E, Zaidi S, Modugno F, Radolec M. et al. Characterization of latently infected EBV+ antibody-secreting B cells isolated from ovarian tumors and malignant ascites. Front Immunol. 2024;15:1379175
26. Sharonov GV, Serebrovskaya EO, Yuzhakova DV, Britanova OV, Chudakov DM. B cells, plasma cells and antibody repertoires in the tumour microenvironment. Nat Rev Immunol. 2020;20:294-307
27. Sautès-Fridman C, Petitprez F, Calderaro J, Fridman WH. Tertiary lymphoid structures in the era of cancer immunotherapy. Nat Rev Cancer. 2019;19:307-25
28. Minici C, Rigamonti E, Lanzillotta M, Monno A, Rovati L, Maehara T. et al. B lymphocytes contribute to stromal reaction in pancreatic ductal adenocarcinoma. Oncoimmunology. 2020;9:1794359
29. Petitprez F, de Reyniès A, Keung EZ, Chen TW-W, Sun C-M, Calderaro J. et al. B cells are associated with survival and immunotherapy response in sarcoma. Nature. 2020;577:556-60
30. Liu R, Lu Z, Gu J, Liu J, Huang E, Liu X. et al. MicroRNAs 15A and 16-1 Activate signaling pathways that mediate chemotaxis of immune regulatory B cells to colorectal tumors. Gastroenterology. 2018;154:637-51
31. Shi J-Y, Gao Q, Wang Z-C, Zhou J, Wang X-Y, Min Z-H. et al. Margin-infiltrating CD20(+) B cells display an atypical memory phenotype and correlate with favorable prognosis in hepatocellular carcinoma. Clin Cancer Res. 2013;19:5994-6005
32. Yang Y, Chen X, Pan J, Ning H, Zhang Y, Bo Y. et al. Pan-cancer single-cell dissection reveals phenotypically distinct B cell subtypes. Cell. 2024;187:4790-811
33. Mizoguchi A, Mizoguchi E, Takedatsu H, Blumberg RS, Bhan AK. Chronic intestinal inflammatory condition generates IL-10-producing regulatory B cell subset characterized by CD1d upregulation. Immunity. 2002;16:219-30
34. Wang WW, Yuan XL, Chen H, Xie GH, Ma YH, Zheng YX. et al. CD19+CD24hiCD38hiBregs involved in downregulate helper T cells and upregulate regulatory T cells in gastric cancer. Oncotarget. 2015;6:33486-99
35. Mirlekar B, Michaud D, Lee SJ, Kren NP, Harris C, Greene K. et al. B cell-derived IL35 drives STAT3-dependent CD8+ T-cell exclusion in pancreatic cancer. Cancer Immunol Res. 2020;8:292-308
36. Mao H, Pan F, Wu Z, Wang Z, Zhou Y, Zhang P. et al. Colorectal tumors are enriched with regulatory plasmablasts with capacity in suppressing T cell inflammation. Int Immunopharmacol. 2017;49:95-101
37. Pylayeva-Gupta Y, Das S, Handler JS, Hajdu CH, Coffre M, Koralov SB. et al. IL35-producing B cells promote the development of pancreatic neoplasia. Cancer Discov. 2016;6:247-55
38. Kobayashi T, Oishi K, Okamura A, Maeda S, Komuro A, Hamaguchi Y. et al. Regulatory B1a cells suppress melanoma tumor immunity via IL-10 production and inhibiting T helper type 1 cytokine production in tumor-infiltrating CD8+ T cells. J Invest Dermatol. 2019;139:1535-44
39. Takahashi R, Macchini M, Sunagawa M, Jiang Z, Tanaka T, Valenti G. et al. Interleukin-1β-induced pancreatitis promotes pancreatic ductal adenocarcinoma via B lymphocyte-mediated immune suppression. Gut. 2021;70:330-41
40. Xie Y, Huang Y, Li Z-Y, Jiang W, Shi N-X, Lu Y. et al. Interleukin-21 receptor signaling promotes metabolic dysfunction-associated steatohepatitis-driven hepatocellular carcinoma by inducing immunosuppressive IgA+ B cells. Mol Cancer. 2024;23:95
41. Lindner S, Dahlke K, Sontheimer K, Hagn M, Kaltenmeier C, Barth TFE. et al. Interleukin 21-induced granzyme B-expressing B cells infiltrate tumors and regulate T cells. Cancer Res. 2013;73:2468-79
42. Mirlekar B, Wang Y, Li S, Zhou M, Entwistle S, De Buysscher T. et al. Balance between immunoregulatory B cells and plasma cells drives pancreatic tumor immunity. Cell Rep Med. 2022;3:100744
43. Lu Z, Liu R, Wang Y, Jiao M, Li Z, Wang Z. et al. Ten-eleven translocation-2 inactivation restrains IL-10-producing regulatory B cells to enable antitumor immunity in hepatocellular carcinoma. Hepatology. 2023;77:745-59
44. Han S, Feng S, Ren M, Ma E, Wang X, Xu L. et al. Glioma cell-derived placental growth factor induces regulatory B cells. Int J Biochem Cell Biol. 2014;57:63-8
45. Miao B-P, Zhang R-S, Li M, Fu Y-T, Zhao M, Liu Z-G. et al. Nasopharyngeal cancer-derived microRNA-21 promotes immune suppressive B cells. Cell Mol Immunol. 2015;12:750-6
46. Li X, Du H, Zhan S, Liu W, Wang Z, Lan J. et al. The interaction between the soluble programmed death ligand-1 (sPD-L1) and PD-1+ regulator B cells mediates immunosuppression in triple-negative breast cancer. Front Immunol. 2022;13:830606
47. Wang Z, Lu Z, Lin S, Xia J, Zhong Z, Xie Z. et al. Leucine-tRNA-synthetase-2-expressing B cells contribute to colorectal cancer immunoevasion. Immunity. 2022;55:1067-81
48. Tousif S, Wang Y, Jackson J, Hough KP, Strenkowski JG, Athar M. et al. Indoleamine 2, 3-dioxygenase promotes aryl hydrocarbon receptor-dependent differentiation of regulatory B cells in lung cancer. Front Immunol. 2021;12:747780
49. Wejksza K, Lee-Chang C, Bodogai M, Bonzo J, Gonzalez FJ, Lehrmann E. et al. Cancer-produced metabolites of 5-lipoxygenase induce tumor-evoked regulatory B cells via peroxisome proliferator-activated receptor α. J Immunol. 2013;190:2575-84
50. Xiao X, Lao X-M, Chen M-M, Liu R-X, Wei Y, Ouyang F-Z. et al. PD-1hi identifies a novel regulatory B-cell population in human hepatoma that promotes disease progression. Cancer Discov. 2016;6:546-59
51. Mao Y, Wang Y, Dong L, Zhang Q, Wang C, Zhang Y. et al. Circulating exosomes from esophageal squamous cell carcinoma mediate the generation of B10 and PD-1high Breg cells. Cancer Sci. 2019;110:2700-10
52. Lee-Chang C, Rashidi A, Miska J, Zhang P, Pituch KC, Hou D. et al. Myeloid-derived suppressive cells promote B cell-mediated immunosuppression via transfer of PD-L1 in glioblastoma. Cancer Immunol Res. 2019;7:1928-43
53. Ye L, Zhang Q, Cheng Y, Chen X, Wang G, Shi M. et al. Tumor-derived exosomal HMGB1 fosters hepatocellular carcinoma immune evasion by promoting TIM-1+ regulatory B cell expansion. J Immunother Cancer. 2018;6:145
54. Das S, Bar-Sagi D. BTK signaling drives CD1dhiCD5+ regulatory B-cell differentiation to promote pancreatic carcinogenesis. Oncogene. 2019;38:3316-24
55. Michaud D, Mirlekar B, Steward C, Bishop G, Pylayeva-Gupta Y. B cell receptor signaling and protein kinase D2 support regulatory B cell function in pancreatic cancer. Front Immunol. 2021;12:745873
56. Shankar S, Stolp J, Juvet SC, Beckett J, Macklin PS, Issa F. et al. Ex vivo-expanded human CD19+TIM-1+ regulatory B cells suppress immune responses in vivo and are dependent upon the TIM-1/STAT3 axis. Nat Commun. 2022;13:3121
57. Li S, Mirlekar B, Johnson BM, Brickey WJ, Wrobel JA, Yang N. et al. STING-induced regulatory B cells compromise NK function in cancer immunity. Nature. 2022;610:373-80
58. Shalapour S, Lin X-J, Bastian IN, Brain J, Burt AD, Aksenov AA. et al. Inflammation-induced IgA+ cells dismantle anti-liver cancer immunity. Nature. 2017;551:340-5
59. Fridman WH, Meylan M, Petitprez F, Sun CM, Italiano A, Sautes-Fridman C. B cells and tertiary lymphoid structures as determinants of tumour immune contexture and clinical outcome. Nat Rev Clin Oncol. 2022;19:441-57
60. Biswas S, Mandal G, Payne KK, Anadon CM, Gatenbee CD, Chaurio RA. et al. IgA transcytosis and antigen recognition govern ovarian cancer immunity. Nature. 2021;591:464-70
61. Garaud S, Zayakin P, Buisseret L, Rulle U, Silina K, de Wind A. et al. Antigen specificity and clinical significance of IgG and IgA autoantibodies produced in situ by tumor-infiltrating B cells in breast cancer. Front Immunol. 2018;9:2660
62. Lin C-W, Xie J, Zhang D, Han KH, Grande G, Wu NC. et al. Immunity against cancer cells may promote their proliferation and metastasis. Proc Natl Acad Sci U S A. 2020;117:426-31
63. Bai Q, Li R, He X, Hong X, Yan Y, Zhao Z. et al. Single-cell landscape of immune cells during the progression from HBV infection to HBV cirrhosis and HBV-associated hepatocellular carcinoma. Front Immunol. 2023;14:1320414
64. Kunkel EJ, Butcher EC. Plasma-cell homing. Nat Rev Immunol. 2003;3:822-9
65. Liu J, Li Y, Lu Z, Gu J, Liang Y, Huang E. et al. Deceleration of glycometabolism impedes IgG-producing B-cell-mediated tumor elimination by targeting SATB1. Immunology. 2019;156:56-68
66. Wei Y, Lao X-M, Xiao X, Wang X-Y, Wu Z-J, Zeng Q-H. et al. Plasma cell polarization to the immunoglobulin G phenotype in hepatocellular carcinomas involves epigenetic alterations and promotes hepatoma progression in mice. Gastroenterology. 2019;156:1890-904
67. Gu Y, Liu Y, Fu L, Zhai L, Zhu J, Han Y. et al. Tumor-educated B cells selectively promote breast cancer lymph node metastasis by HSPA4-targeting IgG. Nat Med. 2019;25:312-22
68. Shalapour S, Font-Burgada J, Di Caro G, Zhong Z, Sanchez-Lopez E, Dhar D. et al. Immunosuppressive plasma cells impede T-cell-dependent immunogenic chemotherapy. Nature. 2015;521:94-8
69. Inoue T, Kurosaki T. Memory B cells. Nat Rev Immunol. 2024;24:5-17
70. Dai D, Gu S, Han X, Ding H, Jiang Y, Zhang X. et al. The transcription factor ZEB2 drives the formation of age-associated B cells. Science. 2024;383:413-21
71. Yolmo P, Rahimi S, Chenard S, Conseil G, Jenkins D, Sachdeva K. et al. Atypical B cells promote cancer progression and poor response to Bacillus Calmette-Guérin in non-muscle invasive bladder cancer. Cancer Immunol Res. 2024;12:1320-39
72. Belderbos RA, Corneth OBJ, Dumoulin D, Hendriks RW, Aerts JGJV, Willemsen M. Atypical B cells (CD21-CD27-IgD-) correlate with lack of response to checkpoint inhibitor therapy in NSCLC. Eur J Cancer. 2024;196:113428
73. Azizi A, Mehdipour F, Samadi M, Rasolmali R, Talei A-R, Ghaderi A. Atypical memory B cells increase in the peripheral blood of patients with breast cancer regardless of lymph node involvement. BMC Immunol. 2024;25:25
74. Kroeger DR, Milne K, Nelson BH. Tumor-infiltrating plasma cells are associated with tertiary lymphoid structures, cytolytic T-cell responses, and superior prognosis in ovarian cancer. Clin Cancer Res. 2016;22:3005-15
75. Garnelo M, Tan A, Her Z, Yeong J, Lim CJ, Chen J. et al. Interaction between tumour-infiltrating B cells and T cells controls the progression of hepatocellular carcinoma. Gut. 2017;66:342-51
76. Ishigami E, Sakakibara M, Sakakibara J, Masuda T, Fujimoto H, Hayama S. et al. Coexistence of regulatory B cells and regulatory T cells in tumor-infiltrating lymphocyte aggregates is a prognostic factor in patients with breast cancer. Breast Cancer. 2019;26:180-9
77. Cho J-W, Park S, Kim G, Han H, Shim HS, Shin S. et al. Dysregulation of TFH-B-TRM lymphocyte cooperation is associated with unfavorable anti-PD-1 responses in EGFR-mutant lung cancer. Nat Commun. 2021;12:6068
78. Singh S, Lee N, Pedroza DA, Bado IL, Hamor C, Zhang L. et al. Chemotherapy coupled to macrophage inhibition induces T-cell and B-cell infiltration and durable regression in triple-negative breast cancer. Cancer Res. 2022;82:2281-97
79. Bruno TC, Ebner PJ, Moore BL, Squalls OG, Waugh KA, Eruslanov EB. et al. Antigen-presenting intratumoral B cells affect CD4+ TIL phenotypes in non-small cell lung cancer patients. Cancer Immunol Res. 2017;5:898-907
80. Cui C, Wang J, Fagerberg E, Chen P-M, Connolly KA, Damo M. et al. Neoantigen-driven B cell and CD4 T follicular helper cell collaboration promotes anti-tumor CD8 T cell responses. Cell. 2021;184:6101-18
81. Zhang B, Vogelzang A, Miyajima M, Sugiura Y, Wu Y, Chamoto K. et al. B cell-derived GABA elicits IL-10+ macrophages to limit anti-tumour immunity. Nature. 2021;599:471-6
82. Pucci F, Garris C, Lai CP, Newton A, Pfirschke C, Engblom C. et al. SCS macrophages suppress melanoma by restricting tumor-derived vesicle-B cell interactions. Science. 2016;352:242-6
83. Liu R-X, Wei Y, Zeng Q-H, Chan K-W, Xiao X, Zhao X-Y. et al. Chemokine (C-X-C motif) receptor 3-positive B cells link interleukin-17 inflammation to protumorigenic macrophage polarization in human hepatocellular carcinoma. Hepatology. 2015;62:1779
84. Ouyang F-Z, Wu R-Q, Wei Y, Liu R-X, Yang D, Xiao X. et al. Dendritic cell-elicited B-cell activation fosters immune privilege via IL-10 signals in hepatocellular carcinoma. Nat Commun. 2016;7:13453
85. Xu X, Meng Q, Erben U, Wang P, Glauben R, Kühl AA. et al. Myeloid-derived suppressor cells promote B-cell production of IgA in a TNFR2-dependent manner. Cell Mol Immunol. 2017;14:597-606
86. Bodogai M, Moritoh K, Lee-Chang C, Hollander CM, Sherman-Baust CA, Wersto RP. et al. Immunosuppressive and prometastatic functions of myeloid-derived suppressive cells rely upon education from tumor-associated B cells. Cancer Res. 2015;75:3456-65
87. Shaul ME, Zlotnik A, Tidhar E, Schwartz A, Arpinati L, Kaisar-Iluz N. et al. Tumor-associated neutrophils drive B-cell recruitment and their differentiation to plasma cells. Cancer Immunol Res. 2021;9:811-24
88. Wang S, Wang H, Li C, Liu B, He S, Tu C. Tertiary lymphoid structures in cancer: immune mechanisms and clinical implications. MedComm (2020). 2024;5:e489
89. Munoz-Erazo L, Rhodes JL, Marion VC, Kemp RA. Tertiary lymphoid structures in cancer - considerations for patient prognosis. Cell Mol Immunol. 2020;17:570-5
90. Li R, Berglund A, Zemp L, Dhillon J, Putney R, Kim Y. et al. The 12-CK score: global measurement of tertiary lymphoid structures. Front Immunol. 2021;12:694079
91. Xu Y, Li Z, Yang Y, Li L, Zhou Y, Ouyang J. et al. A CT-based radiomics approach to predict intra-tumoral tertiary lymphoid structures and recurrence of intrahepatic cholangiocarcinoma. Insights Imaging. 2023;14:173
92. Xu Y, Su G-H, Ma D, Xiao Y, Shao Z-M, Jiang Y-Z. Technological advances in cancer immunity: from immunogenomics to single-cell analysis and artificial intelligence. Signal Transduct Target Ther. 2021;6:312
93. Chen Y-P, Yin J-H, Li W-F, Li H-J, Chen D-P, Zhang C-J. et al. Single-cell transcriptomics reveals regulators underlying immune cell diversity and immune subtypes associated with prognosis in nasopharyngeal carcinoma. Cell Res. 2020;30:1024-42
94. Ruffin AT, Cillo AR, Tabib T, Liu A, Onkar S, Kunning SR. et al. B cell signatures and tertiary lymphoid structures contribute to outcome in head and neck squamous cell carcinoma. Nat Commun. 2021;12:3349
95. Zhang S, Wang B, Ma F, Tong F, Yan B, Liu T. et al. Characteristics of B lymphocyte infiltration in HPV+ head and neck squamous cell carcinoma. Cancer Sci. 2021;112:1402-16
96. Hu Q, Hong Y, Qi P, Lu G, Mai X, Xu S. et al. Atlas of breast cancer infiltrated B-lymphocytes revealed by paired single-cell RNA-sequencing and antigen receptor profiling. Nat Commun. 2021;12:2186
97. Ni Z, Xing D, Zhang T, Ding N, Xiang D, Zhao Z. et al. Tumor-infiltrating B cell is associated with the control of progression of gastric cancer. Immunol Res. 2021;69:43-52
98. Zhang Y, Liu G, Zeng Q, Wu W, Lei K, Zhang C. et al. CCL19-producing fibroblasts promote tertiary lymphoid structure formation enhancing anti-tumor IgG response in colorectal cancer liver metastasis. Cancer Cell. 2024;42:1370-85
99. Olbrecht S, Busschaert P, Qian J, Vanderstichele A, Loverix L, Van Gorp T. et al. High-grade serous tubo-ovarian cancer refined with single-cell RNA sequencing: specific cell subtypes influence survival and determine molecular subtype classification. Genome Med. 2021;13:111
100. Weiner AB, Vidotto T, Liu Y, Mendes AA, Salles DC, Faisal FA. et al. Plasma cells are enriched in localized prostate cancer in Black men and are associated with improved outcomes. Nat Commun. 2021;12:935
101. Helmink BA, Reddy SM, Gao J, Zhang S, Basar R, Thakur R. et al. B cells and tertiary lymphoid structures promote immunotherapy response. Nature. 2020;577:549-55
102. Remark R, Alifano M, Cremer I, Lupo A, Dieu-Nosjean M-C, Riquet M. et al. Characteristics and clinical impacts of the immune environments in colorectal and renal cell carcinoma lung metastases: influence of tumor origin. Clin Cancer Res. 2013;19:4079-91
103. Weed DT, Zilio S, McGee C, Marnissi B, Sargi Z, Franzmann E. et al. The tumor immune microenvironment architecture correlates with risk of recurrence in head and neck squamous cell carcinoma. Cancer Res. 2023;83:3886-900
104. Zhao Y-Y, Fan Z, Tao B-R, Du Z-G, Shi Z-F. Density of tertiary lymphoid structures predicts clinical outcome in breast cancer brain metastasis. J Immunother Cancer. 2024;12:e009232
105. Yin Y-X, Ling Y-H, Wei X-L, He C-Y, Wang B-Z, Hu C-F. et al. Impact of mature tertiary lymphoid structures on prognosis and therapeutic response of Epstein-Barr virus-associated gastric cancer patients. Front Immunol. 2022;13:973085
106. Li J, Zhang L, Xing H, Geng Y, Lv S, Luo X. et al. The absence of intra-tumoral tertiary lymphoid structures is associated with a worse prognosis and mTOR signaling activation in hepatocellular carcinoma with liver transplantation: a multicenter retrospective study. Adv Sci (Weinh). 2024;11:e2309348
107. Zou X, Lin X, Cheng H, Chen Y, Wang R, Ma M. et al. Characterization of intratumoral tertiary lymphoid structures in pancreatic ductal adenocarcinoma: cellular properties and prognostic significance. J Immunother Cancer. 2023;11:e006698
108. Mao Y, Wang X, Xi L, Dong M, Song P, Miao J. et al. Prediction values of tertiary lymphoid structures in the prognosis of patients with left- and right-sided colon cancer: a multicenter propensity score-matched study. Int J Surg. 2023;109:2344-58
109. Ma G, Jia H, Zhang G, Liang Y, Dong X, Fu G. et al. Presence, subtypes, and prognostic significance of tertiary lymphoid structures in urothelial carcinoma of the bladder. Oncologist. 2024;29:e248-e58
110. Sawada J, Hiraoka N, Qi R, Jiang L, Fournier-Goss AE, Yoshida M. et al. Molecular signature of tumor-associated high endothelial venules that can predict breast cancer survival. Cancer Immunol Res. 2022;10:468-81
111. Zhang C, Wang X-Y, Zuo J-L, Wang X-F, Feng X-W, Zhang B. et al. Localization and density of tertiary lymphoid structures associate with molecular subtype and clinical outcome in colorectal cancer liver metastases. J Immunother Cancer. 2023;11:e006425
112. Calderaro J, Petitprez F, Becht E, Laurent A, Hirsch TZ, Rousseau B. et al. Intra-tumoral tertiary lymphoid structures are associated with a low risk of early recurrence of hepatocellular carcinoma. J Hepatol. 2019;70:58-65
113. Li H, Wang J, Liu H, Lan T, Xu L, Wang G. et al. Existence of intratumoral tertiary lymphoid structures is associated with immune cells infiltration and predicts better prognosis in early-stage hepatocellular carcinoma. Aging (Albany NY). 2020;12:3451-72
114. Liu X, Tsang JYS, Hlaing T, Hu J, Ni Y-B, Chan SK. et al. Distinct tertiary lymphoid structure associations and their prognostic relevance in HER2 positive and negative breast cancers. Oncologist. 2017;22:1316-24
115. Lin X, Zhao X, Chen Y, Yang R, Dai Z, Li W. et al. CXC ligand 13 orchestrates an immunoactive microenvironment and enhances immunotherapy response in head and neck squamous cell carcinoma. Int J Immunopathol Pharmacol. 2024;38:3946320241227312
116. Mirlekar B, Michaud D, Searcy R, Greene K, Pylayeva-Gupta Y. IL35 hinders endogenous antitumor T-cell immunity and responsiveness to immunotherapy in pancreatic cancer. Cancer Immunol Res. 2018;6:1014-24
117. Sadeghirad H, Monkman J, Tan CW, Liu N, Yunis J, Donovan ML. et al. Spatial dynamics of tertiary lymphoid aggregates in head and neck cancer: insights into immunotherapy response. J Transl Med. 2024;22:677
118. Patil NS, Nabet BY, Müller S, Koeppen H, Zou W, Giltnane J. et al. Intratumoral plasma cells predict outcomes to PD-L1 blockade in non-small cell lung cancer. Cancer Cell. 2022;40:289-300.e4
119. Kasikova L, Rakova J, Hensler M, Lanickova T, Tomankova J, Pasulka J. et al. Tertiary lymphoid structures and B cells determine clinically relevant T cell phenotypes in ovarian cancer. Nat Commun. 2024;15:2528
120. Hu C, You W, Kong D, Huang Y, Lu J, Zhao M. et al. Tertiary lymphoid structure-associated B cells enhance CXCL13+CD103+CD8+ tissue-resident memory T-cell response to programmed cell death protein 1 blockade in cancer immunotherapy. Gastroenterology. 2024;166:1069-84
121. Ng KW, Boumelha J, Enfield KSS, Almagro J, Cha H, Pich O. et al. Antibodies against endogenous retroviruses promote lung cancer immunotherapy. Nature. 2023;616:563-73
122. Zhang Y, Chen H, Mo H, Hu X, Gao R, Zhao Y. et al. Single-cell analyses reveal key immune cell subsets associated with response to PD-L1 blockade in triple-negative breast cancer. Cancer Cell. 2021;39:1578-93
123. Wang XQ, Danenberg E, Huang C-S, Egle D, Callari M, Bermejo B. et al. Spatial predictors of immunotherapy response in triple-negative breast cancer. Nature. 2023;621:868-76
124. Bertucci F, De Nonneville A, Finetti P, Mamessier E. Predictive power of tertiary lymphoid structure signature for neoadjuvant chemotherapy response and immunotherapy benefit in HER2-negative breast cancer. Cancer Commun (Lond). 2023;43:943-6
125. Gao J, Navai N, Alhalabi O, Siefker-Radtke A, Campbell MT, Tidwell RS. et al. Neoadjuvant PD-L1 plus CTLA-4 blockade in patients with cisplatin-ineligible operable high-risk urothelial carcinoma. Nat Med. 2020;26:1845-51
126. Italiano A, Bessede A, Pulido M, Bompas E, Piperno-Neumann S, Chevreau C. et al. Pembrolizumab in soft-tissue sarcomas with tertiary lymphoid structures: a phase 2 PEMBROSARC trial cohort. Nat Med. 2022;28:1199-206
127. van de Walle T, Vaccaro A, Ramachandran M, Pietilä I, Essand M, Dimberg A. Tertiary lymphoid structures in the central nervous system: implications for glioblastoma. Front Immunol. 2021;12:724739
128. González-Tablas Pimenta M, Otero Á, Arandia Guzman DA, Pascual-Argente D, Ruíz Martín L, Sousa-Casasnovas P. et al. Tumor cell and immune cell profiles in primary human glioblastoma: Impact on patient outcome. Brain Pathology. 2021;31:365-80
129. Hou D, Wan H, Katz JL, Wang S, Castro BA, Vazquez-Cervantes GI. et al. Antigen-presenting B cells promote TCF-1+ PD1- stem-like CD8+ T-cell proliferation in glioblastoma. Front Immunol. 2024;14:1295218
130. Hou D, Castro B, Dapash M, Zolp A, Katz J, Arrieta V. et al. B-cells drive response to PD-1 blockade in glioblastoma upon neutralization of TGFbeta-mediated immunosuppression. Res Sq. 2023;9:2399170
131. Mauldin IS, Jo J, Wages NA, Yogendran LV, Mahmutovic A, Young SJ. et al. Proliferating CD8+ T cell infiltrates are associated with improved survival in glioblastoma. Cells. 2021;10:3378
132. Zhang Y, Jiang C, Trudeau SJ, Narita Y, Zhao B, Teng M. et al. Histone loaders CAF1 and HIRA restrict Epstein-Barr Virus B-cell lytic reactivation. mBio. 2020;11:e01063-20
133. Tempera I, De Leo A, Kossenkov AV, Cesaroni M, Song H, Dawany N. et al. Identification of MEF2B, EBF1, and IL6R as direct gene targets of Epstein-Barr Virus (EBV) nuclear antigen 1 critical for EBV-infected B-lymphocyte survival. J Virol. 2016;90:345-55
134. Chung MKY, Gong L, Kwong DLW, Lee VHF, Lee AWM, Guan XY. et al. Functions of double-negative B cells in autoimmune diseases, infections, and cancers. EMBO Mol Med. 2023;15:e17341
135. Hladíková K, Koucký V, Bouček J, Laco J, Grega M, Hodek M. et al. Tumor-infiltrating B cells affect the progression of oropharyngeal squamous cell carcinoma via cell-to-cell interactions with CD8+ T cells. J Immunother Cancer. 2019;7:261
136. Wang M, Zhai R, Wang M, Zhu W, Zhang J, Yu M. et al. Tertiary lymphoid structures in head and neck squamous cell carcinoma improve prognosis by recruiting CD8+ T cells. Mol Oncol. 2023;17:1514-30
137. Yang J-G, Wang W-M, Xia H-F, Yu Z-L, Li H-M, Ren J-G. et al. Lymphotoxin-α promotes tumor angiogenesis in HNSCC by modulating glycolysis in a PFKFB3-dependent manner. Int J Cancer. 2019;145:1358-70
138. Jeske SS, Brand M, Ziebart A, Laban S, Doescher J, Greve J. et al. Adenosine-producing regulatory B cells in head and neck cancer. Cancer Immunol Immunother. 2020;69:1205-16
139. Molostvov G, Gachechiladze M, Shaaban AM, Hayward S, Dean I, Dias IHK. et al. Tspan6 stimulates the chemoattractive potential of breast cancer cells for B cells in an EV- and LXR-dependent manner. Cell Rep. 2023;42:112207
140. Zou Y, Ye F, Kong Y, Hu X, Deng X, Xie J. et al. The single-cell landscape of intratumoral heterogeneity and the immunosuppressive microenvironment in liver and brain metastases of breast cancer. Adv Sci (Weinh). 2023;10:e2203699
141. Sammut S-J, Galson JD, Minter R, Sun B, Chin S-F, De Mattos-Arruda L. et al. Predictability of B cell clonal persistence and immunosurveillance in breast cancer. Nat Immunol. 2024;25:916-24
142. Tacconi C, Commerford CD, Dieterich LC, Schwager S, He Y, Ikenberg K. et al. CD169+ lymph node macrophages have protective functions in mouse breast cancer metastasis. Cell Rep. 2021;35:108993
143. Huang H, Yao Y, Shen L, Jiang J, Zhang T, Xiong J. et al. CD24hiCD27+ Bregs within metastatic lymph nodes promote multidrug resistance in breast cancer. Clinical Cancer Res. 2023;29:5227-43
144. Mehdipour F, Razmkhah M, Faghih Z, Bagheri M, Talei A-R, Ghaderi A. The significance of cytokine-producing B cells in breast tumor-draining lymph nodes. Cell Oncol (Dordr). 2019;42:381-95
145. Shu L, Tang J, Liu S, Tao Y. Plasma cell signatures predict prognosis and treatment efficacy for lung adenocarcinoma. Cell Oncol (Dordr). 2024;47:555-71
146. Kurebayashi Y, Emoto K, Hayashi Y, Kamiyama I, Ohtsuka T, Asamura H. et al. Comprehensive immune profiling of lung adenocarcinomas reveals four immunosubtypes with plasma cell subtype a negative indicator. Cancer Immunol Res. 2016;4:234-47
147. Vanhersecke L, Brunet M, Guégan J-P, Rey C, Bougouin A, Cousin S. et al. Mature tertiary lymphoid structures predict immune checkpoint inhibitor efficacy in solid tumors independently of PD-L1 expression. Nature Cancer. 2021;2:794-802
148. Chen J, Tan Y, Sun F, Hou L, Zhang C, Ge T. et al. Single-cell transcriptome and antigen-immunoglobin analysis reveals the diversity of B cells in non-small cell lung cancer. Genome Biol. 2020;21:152
149. J Z, Z M, D Z, W W, F M, X W. Enhanced frequency and potential mechanism of B regulatory cells in patients with lung cancer. J Transl Med. 2014;12:304
150. Centuori SM, Gomes CJ, Kim SS, Putnam CW, Larsen BT, Garland LL. et al. Double-negative (CD27-IgD-) B cells are expanded in NSCLC and inversely correlate with affinity-matured B cell populations. J Transl Med. 2018;16:30
151. Bessede A, Peyraud F, Le Moulec S, Cousin S, Cabart M, Chomy F. et al. Upregulation of indoleamine 2,3-dioxygenase 1 in tumor cells and tertiary lymphoid structures is a hallmark of inflamed non-small cell lung cancer. Clin Cancer Res. 2023;29:4883-93
152. Di Modugno F, Di Carlo A, Spada S, Palermo B, D'Ambrosio L, D'Andrea D. et al. Tumoral and stromal hMENA isoforms impact tertiary lymphoid structure localization in lung cancer and predict immune checkpoint blockade response in patients with cancer. EBioMedicine. 2024;101:105003
153. Yamakoshi Y, Tanaka H, Sakimura C, Deguchi S, Mori T, Tamura T. et al. Immunological potential of tertiary lymphoid structures surrounding the primary tumor in gastric cancer. Int J Oncol. 2020;57:171-82
154. Katoh H, Komura D, Konishi H, Suzuki R, Yamamoto A, Kakiuchi M. et al. Immunogenetic profiling for gastric cancers identifies sulfated glycosaminoglycans as major and functional B cell antigens in human malignancies. Cell Rep. 2017;20:1073-87
155. Mori T, Tanaka H, Suzuki S, Deguchi S, Yamakoshi Y, Yoshii M. et al. Tertiary lymphoid structures show infiltration of effective tumor-resident T cells in gastric cancer. Cancer Sci. 2021;112:1746-57
156. Hu H-T, Ai X, Lu M, Song Z, Li H. Characterization of intratumoral and circulating IL-10-producing B cells in gastric cancer. Exp Cell Res. 2019;384:111652
157. Murakami Y, Saito H, Shimizu S, Kono Y, Shishido Y, Miyatani K. et al. Increased regulatory B cells are involved in immune evasion in patients with gastric cancer. Sci Rep. 2019;9:13083
158. Kurebayashi Y, Sugimoto K, Tsujikawa H, Matsuda K, Nomura R, Ueno A. et al. Spatial dynamics of T and B cell response predicts clinical outcome of resectable and unresectable hepatocellular carcinoma. Clin Cancer Res. 2024;30:5230
159. Job S, Rapoud D, Dos Santos A, Gonzalez P, Desterke C, Pascal G. et al. Identification of four immune subtypes characterized by distinct composition and functions of tumor microenvironment in intrahepatic cholangiocarcinoma. Hepatology. 2020;72:965-81
160. Wennhold K, Thelen M, Lehmann J, Schran S, Preugszat E, Garcia-Marquez M. et al. CD86+ antigen-presenting B cells are increased in cancer, localize in tertiary lymphoid structures, and induce specific T-cell responses. Cancer Immunol Res. 2021;9:1098-108
161. Weinstein AM, Giraldo NA, Petitprez F, Julie C, Lacroix L, Peschaud F. et al. Association of IL-36γ with tertiary lymphoid structures and inflammatory immune infiltrates in human colorectal cancer. Cancer Immunol Immunother. 2019;68:109-20
162. Senturk ZN, Akdag I, Deniz B, Sayi-Yazgan A. Pancreatic cancer: Emerging field of regulatory B-cell-targeted immunotherapies. Front Immunol. 2023;14:1152551
163. Kinker GS, Vitiello GAF, Diniz AB, Cabral-Piccin MP, Pereira PHB, Carvalho MLR. et al. Mature tertiary lymphoid structures are key niches of tumour-specific immune responses in pancreatic ductal adenocarcinomas. Gut. 2023;72:1927-41
164. Yao M, Preall J, Yeh JTH, Pappin D, Cifani P, Zhao Y. et al. Plasma cells in human pancreatic ductal adenocarcinoma secrete antibodies against self-antigens. JCI Insight. 2023;8:e172449
165. Capello M, Vykoukal JV, Katayama H, Bantis LE, Wang H, Kundnani DL. et al. Exosomes harbor B cell targets in pancreatic adenocarcinoma and exert decoy function against complement-mediated cytotoxicity. Nat Commun. 2019;10:254
166. Zhu H, Xu J, Wang W, Zhang B, Liu J, Liang C. et al. Intratumoral CD38+CD19+B cells associate with poor clinical outcomes and immunosuppression in patients with pancreatic ductal adenocarcinoma. EBioMedicine. 2024;103:105098
167. Lee KE, Spata M, Bayne LJ, Buza EL, Durham AC, Allman D. et al. Hif1a deletion reveals pro-neoplastic function of B cells in pancreatic neoplasia. Cancer Discov. 2016;6:256-69
168. Workel HH, Lubbers JM, Arnold R, Prins TM, van der Vlies P, de Lange K. et al. A transcriptionally distinct CXCL13+CD103+CD8+ T-cell population is associated with B-cell recruitment and neoantigen load in human cancer. Cancer Immunol Res. 2019;7:784-96
169. Mazor RD, Nathan N, Gilboa A, Stoler-Barak L, Moss L, Solomonov I. et al. Tumor-reactive antibodies evolve from non-binding and autoreactive precursors. Cell. 2022;185:1208-22
170. Montfort A, Pearce O, Maniati E, Vincent BG, Bixby L, Böhm S. et al. A strong B-cell response is part of the immune landscape in human high-grade serous ovarian metastases. Clin Cancer Res. 2017;23:250-62
171. Yang Z, Wang W, Zhao L, Wang X, Gimple RC, Xu L. et al. Plasma cells shape the mesenchymal identity of ovarian cancers through transfer of exosome-derived microRNAs. Sci Adv. 2021;7:eabb0737
172. Wei X, Jin Y, Tian Y, Zhang H, Wu J, Lu W. et al. Regulatory B cells contribute to the impaired antitumor immunity in ovarian cancer patients. Tumour Biol. 2016;37:6581-8
173. Maslanka J, Torres G, Londregan J, Goldman N, Silberman D, Somerville J. et al. Loss of B1 and marginal zone B cells during ovarian cancer. Cell Immunol. 2024;395-396:104788
174. Kobayashi T, Hamaguchi Y, Hasegawa M, Fujimoto M, Takehara K, Matsushita T. B cells promote tumor immunity against B16F10 melanoma. Am J Pathol. 2014;184:3120-9
175. Griss J, Bauer W, Wagner C, Simon M, Chen M, Grabmeier-Pfistershammer K. et al. B cells sustain inflammation and predict response to immune checkpoint blockade in human melanoma. Nat Commun. 2019;10:4186
176. Cabrita R, Lauss M, Sanna A, Donia M, Skaarup Larsen M, Mitra S. et al. Tertiary lymphoid structures improve immunotherapy and survival in melanoma. Nature. 2020;577:561-5
177. Shibad V, Bootwala A, Mao C, Bader H, Vo H, Landesman-Bollag E. et al. L2pB1 cells contribute to tumor growth inhibition. Front Immunol. 2021;12:722451
178. Bekkat F, Seradj M, Lengagne R, Fiore F, Kato M, Lucas B. et al. Upregulation of IL4-induced gene 1 enzyme by B2 cells during melanoma progression impairs their antitumor properties. Eur J Immunol. 2024;54:e2350615
179. Bod L, Kye Y-C, Shi J, Torlai Triglia E, Schnell A, Fessler J. et al. B-cell-specific checkpoint molecules that regulate anti-tumour immunity. Nature. 2023;619:348-56
180. Zhang C, Xin H, Zhang W, Yazaki PJ, Zhang Z, Le K. et al. CD5 binds to interleukin-6 and induces a feed-forward loop with the transcription factor STAT3 in B cells to promote cancer. Immunity. 2016;44:913-23
181. Lee-Chang C, Bodogai M, Moritoh K, Olkhanud PB, Chan AC, Croft M. et al. Accumulation of 4-1BBL+ B cells in the elderly induces the generation of granzyme-B+ CD8+ T cells with potential antitumor activity. Blood. 2014;124:1450-9
182. Masuda T, Tanaka N, Takamatsu K, Hakozaki K, Takahashi R, Anno T. et al. Unique characteristics of tertiary lymphoid structures in kidney clear cell carcinoma: prognostic outcome and comparison with bladder cancer. J Immunother Cancer. 2022;10:e003883
183. Xu W, Lu J, Liu W-R, Anwaier A, Wu Y, Tian X. et al. Heterogeneity in tertiary lymphoid structures predicts distinct prognosis and immune microenvironment characterizations of clear cell renal cell carcinoma. J Immunother Cancer. 2023;11:e006667
184. Wang Y-Q, Chen W-J, Zhou W, Dong K-Q, Zuo L, Xu D. et al. Integrated analysis of tertiary lymphoid structures and immune infiltration in ccRCC microenvironment revealed their clinical significances: a multicenter cohort study. J Immunother Cancer. 2024;12:e008613
185. Li S, Huang C, Hu G, Ma J, Chen Y, Zhang J. et al. Tumor-educated B cells promote renal cancer metastasis via inducing the IL-1β/HIF-2α/Notch1 signals. Cell Death Dis. 2020;11:163
186. Woo JR, Liss MA, Muldong MT, Palazzi K, Strasner A, Ammirante M. et al. Tumor infiltrating B-cells are increased in prostate cancer tissue. J Transl Med. 2014;12:30
187. Tan J, Jin X, Zhao R, Wei X, Liu Y, Kong X. Beneficial effect of T follicular helper cells on antibody class switching of B cells in prostate cancer. Oncol Rep. 2015;33:1512-8
188. Ammirante M, Kuraishy AI, Shalapour S, Strasner A, Ramirez-Sanchez C, Zhang W. et al. An IKKα-E2F1-BMI1 cascade activated by infiltrating B cells controls prostate regeneration and tumor recurrence. Genes Dev. 2013;27:1435-40
189. Piron B, Bastien M, Antier C, Dalla-Torre R, Jamet B, Gastinne T. et al. Immune-related adverse events with bispecific T-cell engager therapy targeting B-cell maturation antigen. Haematologica. 2024;109:357-61
190. Wang S-s, Liu W, Ly D, Xu H, Qu L, Zhang L. Tumor-infiltrating B cells: their role and application in anti-tumor immunity in lung cancer. Cell Mol Immunol. 2019;16:6-18
191. Shen L, Li J, Liu Q, Das M, Song W, Zhang X. et al. Nano-trapping CXCL13 reduces regulatory B cells in tumor microenvironment and inhibits tumor growth. J Control Release. 2022;343:303-13
192. Kar UK, Srivastava MK, Andersson A, Baratelli F, Huang M, Kickhoefer VA. et al. Novel CCL21-vault nanocapsule intratumoral delivery inhibits lung cancer growth. PLoS One. 2011;6:e18758
193. Zhang X, Shen L, Liu Q, Hou L, Huang L. Inhibiting PI3 kinase-γ in both myeloid and plasma cells remodels the suppressive tumor microenvironment in desmoplastic tumors. J Control Release. 2019;309:173-80
194. Aj G, Mm K, T T, Av N, Ni A, B R. et al. Bruton Tyrosine Kinase-dependent immune cell cross-talk drives pancreas cancer. Cancer Discov. 2016;6:270-85
195. Tsuchikama K, Anami Y, Ha SYY, Yamazaki CM. Exploring the next generation of antibody-drug conjugates. Nat Rev Clin Oncol. 2024;21:203-23
196. Wang D, Huang L, Qian D, Cao Y, Wu X, Xu P. et al. Low-dose radiotherapy promotes the formation of tertiary lymphoid structures in lung adenocarcinoma. Front Immunol. 2023;14:1334408
197. Lu Y, Zhao Q, Liao J-Y, Song E, Xia Q, Pan J. et al. Complement Signals Determine Opposite Effects of B Cells in Chemotherapy-Induced Immunity. Cell. 2020;180:1081-97
198. Johansson-Percival A, He B, Li Z-J, Kjellén A, Russell K, Li J. et al. De novo induction of intratumoral lymphoid structures and vessel normalization enhances immunotherapy in resistant tumors. Nat Immunol. 2017;18:1207-17
199. Delvecchio FR, Fincham REA, Spear S, Clear A, Roy-Luzarraga M, Balkwill FR. et al. Pancreatic cancer chemotherapy is potentiated by induction of tertiary lymphoid structures in mice. Cell Mol Gastroenterol Hepatol. 2021;12:1543-65
200. Rodriguez AB, Peske JD, Woods AN, Leick KM, Mauldin IS, Meneveau MO. et al. Immune mechanisms orchestrate tertiary lymphoid structures in tumors via cancer-associated fibroblasts. Cell Rep. 2021;36:109422
201. van Hooren L, Vaccaro A, Ramachandran M, Vazaios K, Libard S, van de Walle T. et al. Agonistic CD40 therapy induces tertiary lymphoid structures but impairs responses to checkpoint blockade in glioma. Nat Commun. 2021;12:4127
202. Semmrich M, Marchand J-B, Fend L, Rehn M, Remy C, Holmkvist P. et al. Vectorized Treg-depleting αCTLA-4 elicits antigen cross-presentation and CD8+ T cell immunity to reject 'cold' tumors. J Immunother Cancer. 2022;10:e003488
203. Lutz ER, Wu AA, Bigelow E, Sharma R, Mo G, Soares K. et al. Immunotherapy converts nonimmunogenic pancreatic tumors into immunogenic foci of immune regulation. Cancer Immunol Res. 2014;2:616-31
204. Zhang Y, Xu J, Fei Z, Dai H, Fan Q, Yang Q. et al. 3D printing scaffold vaccine for antitumor immunity. Adv Mater. 2021;33:e2106768
205. Weinstein AM, Chen L, Brzana EA, Patil PR, Taylor JL, Fabian KL. et al. Tbet and IL-36γ cooperate in therapeutic DC-mediated promotion of ectopic lymphoid organogenesis in the tumor microenvironment. Oncoimmunology. 2017;6:e1322238
206. Kobayashi Y, Watanabe T. Gel-trapped lymphorganogenic chemokines trigger artificial tertiary lymphoid organs and mount adaptive immune responses in vivo. Front Immunol. 2016;7:316
207. Jin X-K, Liang J-L, Zhang S-M, Ji P, Huang Q-X, Qin Y-T. et al. Engineering metal-based hydrogel-mediated tertiary lymphoid structure formation via activation of the STING pathway for enhanced immunotherapy. Mater Horiz. 2023;10:4365-79
208. Wen Z, Liu H, Qiao D, Chen H, Li L, Yang Z. et al. Nanovaccines fostering tertiary lymphoid structure to attack mimicry nasopharyngeal carcinoma. ACS Nano. 2023;17:7194-206
209. Goyal G, Prabhala P, Mahajan G, Bausk B, Gilboa T, Xie L. et al. Ectopic lymphoid follicle formation and human seasonal influenza vaccination responses recapitulated in an organ-on-a-chip. Adv Sci (Weinh). 2022;9:e2103241
Author contact
![]() Corresponding authors: Binhao Zhang (binhaozhangtjmu.edu.cn), Junnan Liang (liangjunnantjmu.edu.cn), Bixiang Zhang (bixiangzhangedu.cn).
Corresponding authors: Binhao Zhang (binhaozhangtjmu.edu.cn), Junnan Liang (liangjunnantjmu.edu.cn), Bixiang Zhang (bixiangzhangedu.cn).