Impact Factor
- Issue 14; 2026
- Issue 13; 2026
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Volume 16; 2026
- Advance Articles
- Past Issues
- Cover Images
- Cover Suggestion
- Index & Coverage
- Special Issues
Introduction
Epidermal growth factor receptor
Human epidermal growth factor...
Platelet-derived growth factor...
Hepatocyte growth factor receptor
Insulin-like growth factor-1...
Insulin-like growth factor-2...
Folate receptor α
Prostaglandin E2 receptor 2
Monocarboxylate transporters
CD146
C-X-C chemokine receptor type 4
Considerations for the...
Conclusion
Abbreviations
Acknowledgements
References
International Journal of Biological Sciences
International Journal of Medical Sciences
 Global reach, higher impact
Global reach, higher impact

Theranostics 2021; 11(8):3882-3897. doi:10.7150/thno.55760 This issue Cite
Review
Molecular targets for anticancer therapies in companion animals and humans: what can we learn from each other?
Irati Beltrán Hernández1,2*, Jannes Z. Kromhout2*, Erik Teske3, Wim E. Hennink1, Sebastiaan A. van Nimwegen3, Sabrina Oliveira1,2 ![]()
1. Pharmaceutics, Department of Pharmaceutical Sciences, Faculty of Science, Utrecht University, 3584 CG Utrecht, the Netherlands.
2. Cell Biology, Neurobiology and Biophysics, Department of Biology, Faculty of Science, Utrecht University, 3584 CH Utrecht, the Netherlands.
3. Department of Clinical Sciences, Faculty of Veterinary Medicine, Utrecht University, 3584 CM Utrecht, the Netherlands.
*Equal contributions to this work.
Received 2020-11-11; Accepted 2020-12-29; Published 2021-2-6
Abstract

Despite clinical successes in the treatment of some early stage cancers, it is undeniable that novel and innovative approaches are needed to aid in the fight against cancer. Targeted therapies offer the desirable feature of tumor specificity while sparing healthy tissues, thereby minimizing side effects. However, the success rate of translation of these therapies from the preclinical setting to the clinic is dramatically low, highlighting an important point of necessary improvement in the drug development process in the oncology field. The practice of a comparative oncology approach can address some of the current issues, by introducing companion animals with spontaneous tumors in the linear drug development programs. In this way, animals from the veterinary clinic get access to novel/innovative therapies, otherwise inaccessible, while generating robust data to aid therapy refinement and increase translational success. In this review, we present an overview of targetable membrane proteins expressed in the most well-characterized canine and feline solid cancers, greatly resembling the counterpart human malignancies. We identified particular areas in which a closer collaboration between the human and veterinary clinic would benefit both human and veterinary patients. Considerations and challenges to implement comparative oncology in the development of anticancer targeted therapies are also discussed.
Keywords: targeted therapy, comparative oncology, companion animals, molecular targets
Introduction
Cancer is an important cause of death in humans worldwide, accountable for approximately 10 million deaths every year [1]. The use of surgery, chemo- and radiotherapy has certainly led to remarkable clinical outcomes for some early stage cancers [2], but the overall need for improved outcome is undeniable. Differences in anatomical location, aetiology, and molecular biology of cancers stress the necessity to develop novel therapies and combined approaches that more specifically address the cancer in an individual patient. Expanding the therapeutic arsenal to fight cancer, numerous targeted therapies and immunotherapies have emerged in the last decade [3, 4]. In particular, continuous development in the field of molecular biology and oncogenes raises opportunities for new molecular targeted therapies, e.g. tyrosine kinase inhibitors (TKI), monoclonal antibodies and antibody-drug conjugates targeting cancer cells. These offer selectivity towards cancer cells over normal cells by exploiting genetic alterations in malignant cells, minimizing damage to healthy tissues and improving patient's quality of life [4]. This attractive feature unique to targeted therapies warrants the innovative efforts of researchers to develop new targeted treatments.
In the oncology field, the success rate of translation of a therapy from preclinical to clinical use is as low as 3.4% [5, 6]. This is the lowest success rate among therapeutic disciplines and can be partly explained by the use of (immunocompromised) rodent xenograft models as the main experimental tool in the field, which certainly fail to reproduce most aspects of such an intricate disease as cancer. These models, in addition to the obvious biological differences between species, often miss the tissue-specific environment, host immunity, the capacity to metastasize and development of treatment resistance [6]. It is clear that a more clinically relevant setting is needed to bridge the currently existing gap and lead to more successful therapies. Comparative oncology offers this, being a discipline that integrates the study of spontaneous cancers in companion animals into studies of human oncology. Consequently, a win-win scenario is created in which therapeutic options can arise for both humans and veterinary patients. This field of study exemplifies the concept of “One Medicine”, which recognises the unity of human and veterinary medicine and that both can advance hand in hand [7]. Dogs and cats with naturally occurring tumors and intact immune systems are of extraordinary translational significance [8]. These veterinary patients present a larger genetic variation than inbred mouse strains and are exposed to environmental factors similar to humans, resulting in less clear-cut results during clinical evaluation that better reflect the human situation. In addition, the shorter life span and lack of therapeutic options for companion animals allow the smooth initiation of therapeutic trials and rapid data collection, while still offering a relatively long-term follow-up [8]. Many cancer types in the veterinary clinic are considered as exceptional models of the human counterpart, exhibiting genetic, molecular and clinical similarities [9, 10]. With the revealing of the dog genome in 2005 and, more recently, the cat genome in 2014, the study of spontaneous cancers in these animals has become even more interesting for comparative oncology [11, 12].
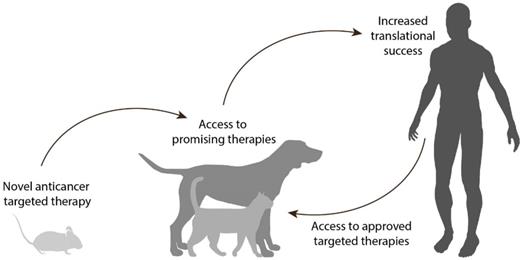
In the context of developing novel targeted therapies, the treatment of veterinary patients with spontaneous tumors can generate robust data and give very valuable insights for treatment refinement, thereby increasing the translational success rate. At the same time, these animals get access to novel and promising therapies, which would otherwise remain inaccessible for these patients (Figure 1).
Proposed approach for the development of anticancer targeted therapies. First, encouraging results are obtained in preclinical studies (e.g. rodents) for a targeted therapy utilizing a particular target. The therapy can be evaluated for the treatment of relevant cancers in companion animals expressing the target in question, allowing access to novel and promising therapeutic approaches for these veterinary patients. Robust data are thus acquired in a more relevant setting, to guide the translation of the therapy towards the equivalent human cancer while increasing translational success. At the same time, the use of current successful targeted therapies in the human clinic can be expanded to the veterinary clinic, based on the knowledge of which targets are present in the different canine and feline cancers.
In this review, we present an overview of molecular targets in companion animals and their expression in relevant cancers (i.e. with translation potential). The focus is on targetable membrane proteins with high homology across species present on the most well-characterized solid cancer models: canine and feline oral squamous cell carcinoma, canine bladder cancer, canine and feline mammary carcinoma, canine osteosarcoma and canine melanoma. A brief introduction of each cancer, its human counterpart and available targeted therapies is given in Table 1, together with a collection of comparative studies highlighting the rationale to focus on the selected cancer types. This table evidences the existing gap between human and veterinary medicine, since no targeted therapies are yet approved for the reviewed cancer types in companion animals. From the listed targeted therapies in Table 1, only the approved monoclonal antibodies cetuximab and trastuzumab are being assessed at the preclinical level for the treatment of the veterinary malignancies, and thus these therapies will be addressed further in this review.
Overview of the translational cancer models included in this review, i.e. human and canine/feline counterpart, available targeted therapies and selected reviews describing similarities
| Cancer | % of all cancers | Approved targeted therapy (target) | Selected review |
|---|---|---|---|
| Human head and neck squamous cell carcinoma | 6% [1] | Cetuximab (EGFR) | [13, 14] |
| Canine and feline oral squamous cell carcinoma | 8 % of all feline cancers [15] 2% of all canine cancers [16] | - | |
| Human invasive transitional cell carcinoma (TCC) | 3% are bladder cancer, of which 20% are invasive TCC [1] | Erdafitinib (FGFR) | [17] |
| Canine invasive TCC | 2% are bladder cancer, of which > 90% are invasive TCC [17] | - | |
| Human mammary carcinoma | 12% (26% in women) [1] | Trastuzumab (HER2), Bevacizumab (VEGF), others | [10, 18] |
| Canine and feline mammary carcinoma | 25% in female dogs [18]; 17% in female cats [19] | - | |
| Human osteosarcoma | 3.5% of all childhood cancers < 1% in adults [20] | Sorafenib (multiple kinases) | [21] |
| Canine osteosarcoma | 2% (especially large size breeds) [22] | - | |
| Human melanoma (non-UV induced) | 6% are melanoma, of which 15 % are non-UV induced [23] | Vemurafenib (BRAF), Trametinib (MEK), others | [24] |
| Canine melanoma | 7% are melanoma, of which > 95% are non-UV induced [25] | - |
EGFR: epidermal growth factor receptor; FGFR: fibroblast growth factor receptor; HER2: human epidermal growth factor receptor 2; VEGF: vascular endothelial growth factor; BRAF: B-Raf proto-oncogene; MEK: mitogen-activated protein kinase.
The different membrane targets that are addressed in this review are presented in Table 2, emphasizing their relevance in the oncology field. Most of these proteins are tyrosine kinase receptors mediating cell proliferation and survival. G protein-coupled receptors and nutrient transporters are also common targets under evaluation in human clinical trials. Table 2 lists some examples of the therapies approved for human use, mostly TKIs (e.g. gefitinib and lapatinib) and the already mentioned monoclonal antibodies (i.e. cetuximab and trastuzumab). Next to these, also other therapies are indicated in Table 2, though only those that were found to be tested in companion animals are described further.
Overview of the membrane targets included in this review and their relevance in oncology
| Membrane target | Ligands/activation | Protein type | Role | Approved therapies (indication) or in trials | Relevance in oncology |
|---|---|---|---|---|---|
| EGFR (ErbB1, HER1) | EGF, TGFα, others | Tyrosine kinase receptor | Cell proliferation; migration; angiogenesis; survival | Cetuximab (colorectal cancer), gefitinib (lung cancer), others | EGFR mutations affecting its expression or activity are common drivers of molecular pathogenesis in many cancers [28] |
| HER2 (ErbB2) | Dimerization with other ErbB family receptors | Trastuzumab and lapatinib (breast and gastric cancer), others | HER2 is an established therapeutic target in a subset of human breast cancers first described in the late 1980s [29] | ||
| PDGFRα/β | PDGF isoforms | Oralatumab (soft tissue sarcoma), imatinib (chronic myeloid leukaemia), others | PDGFRs are among the most commonly inhibited kinases by FDA-approved TKIs to treat cancer [30] | ||
| c-Met (HGFR, Met) | HGF | Cabozantinib (renal cell carcinoma) and crizotinib (lung cancer) | c-Met deregulation correlates with poor prognosis in many human cancers [31] | ||
| IGF-1R | IGF-1, IGF-2 | Xentuzumab (lung, prostate and breast cancer) | When deregulated, IGF-1R plays a role in the pathogenesis of cancer, although it is usually not considered as the main driver [32] | ||
| IGF-2R | IGF-2, mannose-6-phosphate-bearing molecules | Receptor lacking tyrosine kinase activity | Scavenger for circulating IGF-2 | - | Mutations in the M6P/IGF2R gene are frequently found in malignant tumors such as melanoma and mammary carcinoma [33] |
| FRα | Folate | GPI-anchored glycoprotein | Regulate uptake of folate (vitamin B9) | Farletuzumab (ovarian cancer), others | Overexpression of FRα has been associated with several cancers, whereas its expression in normal tissues is limited [34] |
| EP2 | PGE2 | G protein-coupled receptor | Angiogenesis, metastasis, treatment resistance | TPST-1495 (colorectal cancer) | EP2 is abnormally expressed in many cancers, including prostate, bladder and breast cancers [35] |
| MCTs | Lactate, pyruvate ketone bodies, others | Proton-linked channel | Transmembrane transport of monocarboxylate metabolites | AZD3965 (lymphoma) | Due to the high glycolytic activity of tumor cells, MCT1 and MCT4 are highly expressed by aggressive and metastasizing tumors [36] |
| CD146 (MCAM) | Laminin 411, VEGFR2, others | Immunoglobulin superfamily receptor | Adhesion molecule and cell survival | - | In human melanomas, CD146 is largely involved in tumor progression [37] |
| CXCR4 (CD184) | CXC12 | G protein-coupled receptor | Cell proliferation and survival | Ulocuplumab (multiple myeloma) | CXCR4 expression is low or absent in many healthy tissues, but it is expressed in over 20 types of cancer [38] |
EGFR: epidermal growth factor receptor; TGFα: transforming growth factor alpha; HER2: human epidermal growth factor receptor 2; PDGFR-α/β: platelet derived growth factor receptor α/β; c-Met: tyrosine-protein kinase Met; HGFR: hepatocyte growth factor receptor; TKI: tyrosine kinase inhibitor; IGF-1/2R: insulin-like growth factor 1/2 receptor; FRα: folate receptor α; GPI: glycosylphosphatidylinositol; EP2: prostaglandin E2 receptor 2; PGE2: prostaglandin E2; MCT1/4: monocarboxylate transporter 1/4; CD146: cluster of differentiation 146; MCAM: melanoma cell adhesion molecule; VEGFR: vascular endothelial growth factor receptor; CXCR4: C-X-C chemokine receptor type 4; CXC12: C-X-C chemokine 12.
Knowing the targets overexpressed in these relevant animal cancers, not only serves to guide the evaluation of targeted therapies in clinically relevant models of the human disease, but also to expand the use of current successful targeted therapies in humans towards the veterinary clinic. The feasibility of the proposed approach is supported by the fact that cancer remains a leading cause of death not only in humans, but also in companion animals. To compare, dogs develop cancer at a similar rate as humans (1 in 3), while cats are less affected (1 in 4 to 5) [26]. About 50 % of dogs and 30 % of cats over age ten die from cancer. Most of the discussed animal cancers are often compared to adult human cancers, since those are rare during childhood and have a higher incidence in geriatric animals, while canine osteosarcoma is considered to closely resemble the pediatric cancer [27].
Epidermal growth factor receptor
Head and neck squamous cell carcinoma
EGFR overexpression has been reported in up to 90% of human head and neck squamous cell carcinomas (HNSCC) and also in feline oral squamous cell carcinoma (OSCC), making it an interesting therapeutic target in both species [39-41]. Although several EGFR-targeting small molecule inhibitors and monoclonal antibodies are currently approved and under clinical evaluation for the treatment of human HNSCC [42, 43], clinical evaluation of agents targeting EGFR in feline OSCC has not been reported. In vitro studies with a feline OSCC cell line have been conducted to target EGFR with the TKI gefitinib and RNA interference [44, 45]. While gefitinib caused a reduction in cell proliferation, this was accomplished at a relatively high dose and gefitinib resistance developed. By combining siRNA with gefitinib treatment, it was possible to surpass the acquired resistance. Interestingly, the combination of siRNA targeting of EGFR and radiation therapy showed an additive cytotoxic effect in the feline cell line [44]. These results, although scarce and in an initial stage, support the notion of EGFR targeting to treat feline OSCC.
Invasive transitional cell carcinoma
EGFR is overexpressed in both human and canine invasive transitional cell carcinoma (TCC) in about 75% of the cases [17, 46, 47]. Several EGFR-targeting compounds have been or are currently evaluated in clinical trials of human TCC, but so far with mixed results [17]. Recently, a particular 67-gene signature has been associated with sensitivity to EGFR inhibition in human bladder cancer cell lines [48]. Regarding canine invasive TCC, there is only one clinical study involving EGFR-targeting as therapy [49]. In this study, an EGF‐conjugated anthrax toxin was used to target EGFR-expressing human and canine TCC cell lines. After encouraging in vitro results, the effects of the EGF-toxin were evaluated in 6 dogs with invasive TCC, resulting in a 30% reduction of the tumor size in all dogs. These data show promising effects and follow-up studies are warranted. In another recent study, near-infrared photoimmunotherapy was used to target EGFR in canine invasive TCC cell lines [50]. The EGFR-targeting monoclonal antibody can225IgG (caninized cetuximab) was conjugated to a photosensitizer (IRDye700DX) and tested in canine TCC cell lines, showing specificity and cytotoxic potency. Using a xenograft mice model of canine invasive TCC, NIT-PIT treatment led to a statistically significant inhibition of tumor growth and prolonged survival, highlighting thus the promise of the approach.
Osteosarcoma
EGFR is frequently overexpressed in human osteosarcoma (OSA); however, the exact contribution of EGFR in OSA prognosis and development is still not fully understood [51, 52]. EGFR is expressed in canine OSA cell lines and tissues, and a higher expression correlates with a poor prognosis [53, 54]. Currently, several ongoing clinical trials evaluate the use of EGFR-targeting agents in human patients with OSA [52]. As for canine OSA, only in vitro data are available so far [54-56]. Three small molecule EGFR-targeting TKIs, i.e. erbstatin analogue, erlotinib and gefitinib, have been tested alone or in combination with different therapeutic approaches. The three drugs showed activity as monotherapy, but combinations yielded better results. In particular, erbstatin analogue was tested in combination with the chemotherapy agent doxorubicin, resulting in increased cell death compared to erbstatin monotherapy. Erlotinib was tested in combination with radiation therapy and enhanced sensitivity to radiation was observed. Lastly, the combination of gefitinib and the c-Met inhibitor crizotinib showed an additive effect on cell proliferation inhibition. Altogether, combination strategies involving EGFR targeting show potential for the treatment of canine OSA.
Mammary carcinoma
EGFR expression is found in 15-45% of human breast cancer tissues [57]. Accordingly, the EGFR TKIs gefitinib, erlotinib, lapatinib and afatinib are approved for the treatment of some subsets of breast cancer, while the evaluation of more EGFR-targeting drugs is ongoing in clinical trials [58]. In canine mammary carcinoma (CMC) tissues, similar EGFR expression as in the human equivalent cancer has been reported [59]. Similarly, EGFR was found to be significantly overexpressed in cell lines, tissues and xenograft mouse models of feline mammary carcinoma (FMC) [60]. Several studies report cell proliferation inhibition induced by EGFR-targeting in CMC cell lines. Cetuximab and its caninized version (can225IgG), gefitinib and EGFR siRNA have all inhibited cell proliferation as a single agent [61-63]. Only in one of these studies a FMC cell line was included, also showing cell proliferation inhibition after gefitinib treatment [62].
Melanoma
EGFR has also been of interest as a therapeutic target in human melanomas, and it can play a role in the progression of cutaneous melanomas [64]. Two clinical phase II trials have been conducted with EGFR-targeting agents, but the outcomes were poor in human patients with metastatic melanomas [65, 66]. Therefore, there are currently no EGFR-targeted therapies approved for human melanomas. In canine melanoma, little is known regarding EGFR expression and its role. One recent study reported the expression of EGFR and other receptors from the EGFR family in canine melanomas, concluding that EGFR overexpression occurs only in 5% of the oral melanomas and 16% of the skin melanomas [67]. In addition, an increased proliferative index was associated with a decreased EGFR expression in the skin melanomas. In another recent study, the expression of EGFR was found downregulated in oral melanoma compared to the healthy oral tissue [68]. Therefore, EGFR does not seem to be a promising target in human and canine melanoma, but targeting of this receptor in the canine disease has not been investigated yet.
Human epidermal growth factor receptor 2
Invasive transitional cell carcinoma
HER2 overexpression is common in human TCC and it is, therefore, an interesting target [69]. Likewise, significant overexpression of HER2 compared to normal canine bladder has been described in canine invasive TCC tissues at both the RNA and protein level [70-72]. While multiple clinical trials are ongoing to evaluate different HER2-targeting drugs in human patients with TCC [73], the targeting of this protein has not been yet studied in the context of canine TCC. The outcome of the current human clinical trials will certainly shed light on the value of targeting HER2 in this type of bladder cancer and may spur research also in the field of veterinary oncology.
Osteosarcoma
The role of HER2 in human OSA has been controversial. Multiple studies report overexpression of HER2, while others describe the opposite [74-76]. As for canine OSA, overexpression of HER2 has been detected in different OSA cell lines and tissues at the RNA level [77]. Despite the controversial findings on HER2 in human OSA, a clinical trial has been conducted with trastuzumab, a HER2-targeting monoclonal antibody, in combination with chemotherapy [78]. The outcome was rather disappointing and HER2 positive patients did not have a significant therapeutic benefit. Nevertheless, HER2 seems to remain an interesting target in these cancers for targeted therapy and, particularly, for immunotherapy [52]. In human patients with HER2-positive OSA, HER2-specific chimeric antigen receptor T cell therapy has shown initial promising results [79]. Furthermore, dogs with HER2-positive OSA were successfully treated with a vaccine consisting of recombinant Listeria bacteria expressing a chimeric human HER2, which induced HER2-specific immunity and reduced the incidence of metastasis [80]. Altogether, it is reasonable to consider immunotherapies involving HER2 targeting as a therapeutic option for both human and canine OSA, but the targeting of this receptor alone has not yet shown promise.
Mammary carcinoma
HER2 is a major marker for human breast cancer classification and its presence or absence mainly guides the choice of therapeutic strategy. HER2 overexpression is found in 10-40% of human breast cancers [81]. For the treatment of HER2-positive breast cancer, multiple HER2-targeting agents are already approved, e.g. trastuzumab, pertuzumab and afatinib, and others are being tested in ongoing clinical trials [58]. In CMC and FMC, HER2 overexpression is in the same range as in human breast cancers [82-85]. Nonetheless, studies of HER2-targeting in these veterinary patients have been so far limited to in vitro evaluations. The anti-HER2 antibody trastuzumab was able to inhibit cell proliferation in two CMC cell lines [61]. On the other hand, using other CMC and FMC cell lines, the use of the HER2 inhibitor AG825 or a HER2 siRNA only caused a slight inhibition of cell proliferation [62]. All things considered, the genetic, molecular and clinical similarities between the human and canine/feline disease support the value of targeting HER2 in companion animals with mammary carcinoma.
Melanoma
The role of HER2 in human melanomas is still under debate, because of contradictory results regarding the overexpression of this target [86-88]. In canine oral and skin melanomas, the role of HER2 is still largely unknown and there is only one study addressing this. HER2 membranous expression was found in 50 and 43% of the oral and skin melanomas, respectively, but this was not considered to be overexpressed [67]. HER2 expression in the oral melanomas was associated with tumor progression, because of its correlation with emboli occurrence. So far, no solid conclusion can be drawn regarding the potential of HER2 targeting to treat melanoma.
Platelet-derived growth factor receptors
Osteosarcoma
Both PDGFRα and PDGFRβ are frequently overexpressed in OSA human tissues [89, 90]. In canine OSA tissue, the expression of both proteins is commonly found and, in canine OSA cell lines, overexpression of the two receptors has been reported at the RNA level [91]. However, clinical trials evaluating different PDGFR-targeted therapies in human OSA have not been successful. As a result, it has been suggested to target PDGFRs not as monotherapy, but rather in combination with other therapeutic agents such as check point inhibitors or chemotherapy [90]. There are no studies available in which the targeting of PDGFRs in canine OSA has been investigated, but combination strategies might be the answer as in the case of the human disease.
Melanoma
In human cutaneous melanomas, overexpression of PDGFRα is associated with decreased cell proliferation, whereas the role of PDGFRβ in melanomas is, on the other hand, less clear [92, 93]. Still, several phase II and III clinical trials are being conducted in patients with malignant melanomas evaluating TKIs that target PDGFRs amongst other tyrosine kinase receptors [94]. In canine oral melanomas, the co-expression of PDGFRα and PDGFRβ has been described in 37.5% of the cases, which corresponded with a significant shorter overall survival compared to dogs without co-expression [95]. Although the targeting of exclusively PDGFRs has not been investigated yet in companion animals with melanoma, the encouraging results from human clinical trials may instigate researchers in this direction.
Hepatocyte growth factor receptor
Osteosarcoma
c-Met is frequently found overexpressed in human and canine OSA tissues and cell lines [54, 96-98]. Nevertheless, to date, there are no clinical data available of c-Met-targeting agents to treat OSA. The two approved multi-TKI cabozantinib and crizotinib, which target c-Met amongst others, have been tested in human OSA cell lines and xenograft mouse models with promising results [99, 100]. Liao et al. tested a c-Met-targeting drug called PF2362376 in canine OSA cell lines, which inhibited c-Met and induced cell death at a high dose [101]. Another drug that targets c-Met, called PHA‐665752, and a c-Met shRNA have also been evaluated as single agents in two canine OSA cell lines. Although the two agents did not affect cell proliferation, the motility of the cell lines was decreased [98]. Although not clear yet whether targeting c-Met alone can be of value, there are indications that c-Met inhibition can be a promising strategy to treat OSA when also other oncogenic pathways are being targeted.
Insulin-like growth factor-1 receptor
Osteosarcoma
The role of IGF-1 and IGF-1R has been investigated in several human cancers, including OSA [32]. A significant overexpression of IGF-1R has been found in human OSA tissues compared to healthy bone tissue [102]. In canine OSA cell lines and tissues, high expression of IGF-1R is frequently observed at both the RNA and protein level [103, 104]. One clinical study assessing the efficacy of an IGF-1R-targeting drug has shown poor results in humans with OSA [105]. Nonetheless, more IGF-1R-targeting drugs are being investigated in clinical trials to treat human OSA [106], while no data are available concerning canine OSA. The results of the ongoing human clinical trials, when promising, may guide the use of similar therapeutic strategies in dogs with OSA.
Mammary carcinoma
Expression of IGF-1R is frequently found in human breast cancer with varying percentages in the different cancer subtypes [107]. Therefore, IGF-1R has been an interesting target in breast cancer and multiple clinical trials have taken place with IGF-1R-targeting agents [108]. However, these clinical trials showed poor outcomes, resulting in no approved IGF-1R-targeting drugs in human breast cancer so far. In multiple studies with tissue samples of canine mammary gland carcinomas and invasive CMC, high expression of IGF-1R was commonly found, but contradictory findings have been reported regarding its correlation with prognosis in CMC [109-111]. The role and expression of IGF-1R has not been studied in FMC, and no therapeutic strategies targeting this receptor have been assessed in CMC or FMC. Clearly, while human data discourages IGF-1R targeting in breast cancer, more research is needed to get a better understanding of the potential of this approach in companion animals with mammary carcinoma.
Melanoma
IGF-1R often plays a role in the different types of human melanomas and, therefore, it has been considered an interesting drug target for this disease [112, 113]. Among multiple tyrosine kinase receptors, IGF-1R has been found to be the most abundant in canine malignant melanoma cell lines, and its expression was subsequently confirmed in the majority of canine melanoma tissues [114]. There are several preclinical studies showing promising effects of IGF-1R targeting in combination with other therapeutic agents for the treatment of human melanoma [115]. Nevertheless, the use of IGF-1R-targeting agents has not been evaluated in canine melanomas, neither in vitro nor in vivo. It is reasonable to consider IGF-1R a good target in canine melanoma, based on its high expression and the so far encouraging results in human melanoma.
Insulin-like growth factor-2 receptor
Osteosarcoma
In contrast to IGF-1R, research conducted with IGF-2R as a therapeutic target is still in the initial phases. In human primary OSA cell lines, IGF-2R has been found to be significantly overexpressed [96]. Monoclonal antibodies targeting IGF-2R conjugated to different cytotoxic radioisotopes, for radioimmunotherapy, have been evaluated in human OSA cell lines as well as patient-derived xenograft mouse models [116, 117]. The authors showed specificity of the radiolabelled antibody and the ability to supress tumor growth. Furthermore, the same authors described IGF-2R expression in canine OSA tissue, making it an interesting target for further research [116].
Folate receptor α
Invasive transitional cell carcinoma
Dhawan et al. investigated the expression of FRs in human and canine invasive TCC tissues, which was found in 78 and 76% of the cases, respectively [118]. In the same study, the FR-targeting drug folate-vinblastine conjugate was intravenously administered to treat 9 dogs with FR-positive invasive TCC. The drug led to partial remission in 56% of the dogs and stable disease in 44%. The same research group continued this line of investigation and treated 28 dogs with invasive TCC with a different FR-targeting drug, i.e. folate-tubulysin conjugate. This time, clinical benefit was observed in 71% of the dogs, including partial remission in 11% and stable disease in 60% [119]. The encouraging results justify the assessment of FR-targeting agents to treat invasive TCC in the human clinic as well.
Prostaglandin E2 receptor 2
Invasive transitional cell carcinoma
The expression levels of EP2 in human urothelial cancer tissues have been shown to be significantly upregulated compared to normal urothelium [120]. Interestingly, a decreased EP2 expression in the cytoplasm and nucleus in bladder cancer tissues compared to normal bladder has also been reported [121]. When comparing the gene expression patterns of invasive canine TCC tissues and normal bladder, the gene encoding for EP2, PTGER2, was the most upregulated gene in the TCC samples [72]. After that finding, EP2 protein expression was detected in the tumor cells of 11 out of 15 canine TCC tissues, whereas EP2 was absent in normal epithelial cells. Altogether, the overexpression of EP2 in human and canine TCC makes it a promising target, but the efficacy of targeting agents has yet to be investigated.
Monocarboxylate transporters
Head and neck squamous cell carcinoma
Increased expression of MCT1 and MCT4 in human oral SCC has been correlated with a poor survival prognosis and, therefore, MCT1 and MCT4 were proposed as therapeutic targets in HNSCC [122, 123]. More recently, high expression of MCT1 in feline OSCC cell lines and tissues was reported, whereas MCT4 expression was relatively low [124]. Accordingly, the action of a dual MCT1- and MCT4-inhibiting drug, MD-1, was tested in feline OSCC and human HNSCC cell lines as well as in an orthoptic feline OSCC xenograft mouse model. The targeting agent induced cell death in the human and feline cell lines and inhibited tumor growth in the orthoptic mouse model, highlighting the promise of MCTs as therapeutic targets in both species.
CD146
Melanoma
CD146 is highly expressed in human primary and metastatic melanomas and is a marker of poor prognosis in these patients [37]. Multiple antibodies have been developed to target human CD146 and tested in preclinical settings with promising results [37, 125, 126]. In canine tissues of oral and skin melanoma, CD146 is frequently highly expressed as well, especially in oral melanomas [127, 128]. However, at present no studies have been published that investigated CD146 targeting in canine melanoma. Clinical trials to be conducted in the near future will give insights into the value of CD146 as a therapeutic target in the clinical setting.
C-X-C chemokine receptor type 4
Mammary carcinoma
Targeting CXCR4 in human breast cancer patients is currently under evaluation in clinical trials [129]. In FMC tissues, CXCR4 is frequently highly expressed, whereas CXCR4 expression is not detected in healthy mammary tissues of cats [130, 131]. Moreover, CXCR4 has been described as one of the top upstream regulators in canine mammary tumors [132]. Further research in companion animals may be encouraged by the outcome of the current clinical studies in human patients [129].
Multi-kinase inhibitors
Many targetable proteins are tyrosine kinase receptors on the cell membrane. Upon their activation (e.g. ligand binding), intracellular signaling occurs, inducing a multitude of cellular processes, including cell proliferation, differentiation and survival. Alteration in expression and/or activation of tyrosine kinase receptors is common in many human cancers and, therefore, these proteins have been of interest in the field of targeted therapeutics [133].
Sunitinib (Sutent®) and toceranib phosphate (Palladia®) are two drugs targeting multiple of these tyrosine kinase receptors simultaneously, including the vascular endothelial growth factor receptor (VEGFR), PDGFR, c-Kit, colony stimulating factor 1 receptor and fms-like tyrosine kinase 3 [134-136]. Sunitinib and toceranib phosphate are actually structural analogues, used for the treatment of cancer in the human and veterinary clinic, respectively. Sunitinib has been approved for the treatment of different cancers, such as renal cell carcinoma and gastrointestinal stromal tumors. Toceranib phosphate was the first drug approved for targeted cancer treatment in companion animals, indicated for canine mast cell tumors.
Another TKI with a broad range of targets is dasatinib (Sprycel, Dasanix), which targets several tyrosine kinase receptors such as c-Kit and PDGFRs, as well as the SRC kinase family of non-receptor signaling proteins. This drug has already been approved by the FDA for treatment of two types of human leukaemia [137], and its use in the veterinary clinic is gaining interest. Masitinib mesylate (Kinavet, Masivet) is another multi-TKI mainly inhibiting c-Kit and, to a lesser extent, other tyrosine kinase receptors such as PDGFRs and fibroblast growth factor receptor 3 (FGFR3) [138]. Masitinib is approved for the treatment of canine mast cell tumors, while its evaluation in humans with solid tumors, such as pancreatic and colorectal cancer, is ongoing. An overview of the evaluation of toceranib phosphate, dasatinib and masitinib mesylate in the veterinary clinic (for the cancer models here reviewed) is shown in Table 3.
Overview of the use of toceranib phosphate, dasatinib and masitinib mesylate in veterinary clinical studies and in vitro
| Cancer | Toceranib phosphate | Dasatinib | Masitinib mesylate | Ref. |
|---|---|---|---|---|
| HNSCC | Dogs & cats | Canine & feline cells | [134, 135, 139] | |
| TCC | Dogs | [140] | ||
| OSA | Dogs | Dogs | [135, 141, 142] | |
| MC | Dogs | [143, 144] | ||
| MM | Dogs | Dogs | [144, 145] |
HNSCC: head and neck squamous cell carcinoma; TCC: transitional cell carcinoma; OSA: osteosarcoma; MC: mammary carcinoma; MM: malignant melanoma.
Head and neck squamous cell carcinoma
The use of sunitinib has been evaluated in phase II clinical trials in recurrent or metastatic HNSCC, but resulted in poor outcomes due to low efficacy [146, 147]. Since then, toceranib phosphate has been tested in different cancers in dogs and cats, including canine and feline OSCC [134, 135, 148]. London et al. treated 8 dogs with carcinoma in the head and neck region with toceranib phosphate, and 6 of these dogs experienced clinical benefit [135]. In a retrospective study of cats with OSCC, toceranib phosphate treatment significantly extended the lifespan compared to untreated cats [134]. Here the treatment was combined with the administration of non-steroidal anti-inflammatory drugs (NSAIDs) and the beneficial effect of toceranib phosphate alone was not demonstrated. Nevertheless, overall, promising results have been observed so far for the use of toceranib phosphate in the treatment of OSCC in companion animals.
The use of masitinib, which has the highest affinity for c-Kit, has been evaluated in feline and canine OSCC cell lines. Overexpression and activation of c-Kit was found in human HNSCC [149], and similar expression levels have been detected in feline and canine OSCC cell lines [139]. Masitinib inhibits the downstream pathway of c-Kit in these cell lines, but also induces expression of cyclooxygenase 2 (COX2), an enzyme involved in cell proliferation. Consequently, co-treatment with piroxicam (a COX2 inhibitor) resulted in the best inhibition of cell proliferation for the different SCC cell lines [139]. Clearly, further research is still needed to elucidate the promise of these combination strategies.
Invasive transitional cell carcinoma
Sunitinib has been evaluated in several phase II clinical trials of human patients with non-invasive or metastatic TCC. The general outcome of those studies was unsatisfactory, despite clinical benefit in some patients [150-152]. The use of toceranib phosphate has been investigated for the treatment of canine TCC. The expression of PDGFRβ, VEGFR2 and c-Kit, all targets of toceranib phosphate, has been described in canine invasive TCC, although only PDGFR-β was found significantly overexpressed in comparison to normal bladder tissue [153]. Owing to these findings, a retrospective study was recently performed regarding toceranib phosphate treatment in dogs with TCC, showing clinical benefit in 86.7% of the dogs [140]. The study had some limitations, such as the co-administration of NSAIDs which can positively affect overall survival and the presumptive diagnosis of 12 out of 27 dogs. Nonetheless, the results are encouraging for the use of toceranib phosphate to treat canine TCC.
Osteosarcoma
Sunitinib has shown promising effects in human OSA cell lines and xenograft mouse models [154], but no clinical data are available to date. The use of toceranib phosphate in canine OSA has been evaluated in the last couple of years. First, a retrospective study showed the first preliminary evidence of clinical benefits in canine OSA [135]. However, follow-up studies failed to demonstrate a significant clinical benefit of toceranib phosphate as monotherapy or in combination with chemotherapy or other therapeutic agents [155-158]. In a recent paper by Sánchez-Cespedes et al., toceranib phosphate was investigated in canine OSA cell lines and in two orthotopic xenograft canine OSA mouse models, resulting in decreased tumor growth only in one of the two mouse models [159]. The OSA cell line used for this particular xenograft had high expression of PDGFR, c‐Kit and VEGFR2 and, as a result, it has been suggested that toceranib phosphate is more effective in tumors with high expression of these receptors.
Dasatinib has been tested in human clinical trials for the treatment of different sarcomas, including OSA, but there was insufficient activity as a single agent [160]. In a single-case study of a dog with OSA, dasatinib was used as a treatment [141]. The treated dog in the study first underwent surgery, by which primary tumor cells were obtained and cultured. Together with other canine OSA cell lines, the primary cells were screened for 86 small molecule inhibitors. Here dasatinib showed the most promising result and, consequently, was used to treat the dog after receiving surgery and chemotherapy. Eight months after completion of the treatment, the tumor had not recurred. In line with these observations, Marley et al. treated 4 dogs with OSA with dasatinib after surgery and chemotherapy, leading to a prolonged survival time and indicating the potential benefit of this drug for the treatment of canine OSA [142].
Mammary carcinoma
Multiple phase III clinical trials of sunitinib treatment in patients with advanced breast cancer, either as monotherapy or in combination with chemotherapy, have not shown satisfactory results. Therefore, sunitinib has not been approved as treatment in human breast cancer [161-163]. Evaluation of toceranib phosphate in a clinical setting to treat dogs bearing mammary carcinomas has been limited. Five dogs with mammary carcinoma were included and 2 of these had a partial response [144]. Eighteen dogs with inflammatory mammary carcinoma were treated with toceranib phosphate in combination with piroxicam and thalidomide. Of those 18 dogs, 4 received additional radiation therapy, which led to partial response and significant increased survival time compared to the dogs which did not receive radiotherapy. For the 14 dogs not treated with radiation therapy, the combination treatment of toceranib phosphate, piroxicam and thalidomide caused a partial response in 3 dogs and 6 dogs experienced stable disease [143]. Altogether, toceranib phosphate may be effective against canine mammary carcinoma in a multimodal approach, but the effectiveness as a single agent remains to be elucidated.
Melanoma
Sunitinib has been evaluated in human clinical trials to treat cutaneous, mucosal and uveal melanomas. Nevertheless, sunitinib has shown limited activity in those trials and it has been suggested that sunitinib could be more beneficial in combination with other drugs for the treatment of melanoma [164, 165]. In two canine clinical studies evaluating toceranib phosphate, a total of 4 dogs with melanoma were included, from which 3 dogs achieved a stable disease [144, 145]. Although the first results are encouraging, the benefit of toceranib phosphate to treat canine melanomas needs to be addressed in larger studies.
The use of masitinib mesylate has also been evaluated in dogs with melanoma, strongly driven by the fact that c-Kit is a therapeutic target in human melanomas, especially in advanced melanomas with a mutated c-Kit gene [166]. Different c-Kit inhibitors have already been tested in human clinical trials and imatinib is currently recommended for second-line treatment of c-Kit-mutated melanoma [167]. In canine cutaneous and mucosal melanomas, only a few studies have investigated the expression, mutations and role of c-Kit, and the precise role of c-Kit is still not completely understood in these melanomas [168-171]. In a recent study by Giuliano & Dobson, masitinib mesylate was evaluated as a treatment in 17 dogs with advanced malignant melanomas. Only 2 dogs with mucosal melanomas had a partial response, while 7 dogs achieved stable disease. The researchers concluded that masitinib mesylate treatment is not efficient, at least as monotherapy, for advanced malignant melanoma in dogs [172].
Considerations for the development of targeted therapies in comparative oncology
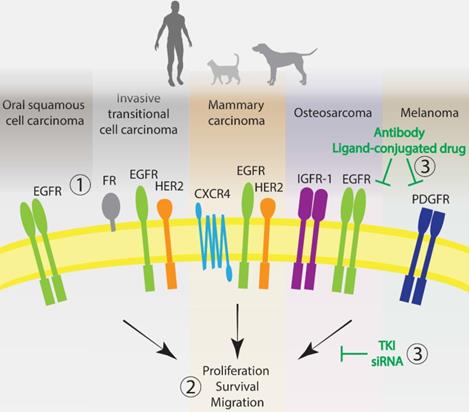
The use of targeted therapies to treat human cancers has been significantly more implemented than in the veterinary clinic [173]. Nevertheless, with the latest developments in molecular biology and the veterinary approval of the two multi-TKIs toceranib and masitinib, targeted therapies in veterinary oncology are certainly becoming a more important focus of cancer research. Dogs with spontaneous cancers have been more of interest in comparative oncology than their feline comrades [10, 174]. However, for some cancer types, cats can be more relevant for translational research; for instance, as a model for human HNSCC and metastatic breast cancer [10]. Most advances in targeted therapies in the veterinary landscape are directed towards canine lymphomas and mast cell tumors. In this review, we have focused on the translation of targeted therapies for solid tumors, by presenting an overview of molecular targets in human cancers and the counterpart malignancy in companion animals, as well as the outcomes of the so far evaluated therapies. To enable relevant and translational research, we focused on targetable membrane proteins with high homology degree between species and their presence in five well-characterized solid cancer types, all considered exceptional models of the respective human disease. Table 4 presents a summary of the reviewed targets, their homology between human and dog or cat, the targeted therapies, and the stage of testing (i.e. cell lines, xenografts in mice, or companion animals). While multi-TKIs are more commonly evaluated in veterinary patients, the targeting of individual proteins is mainly restricted to preclinical research. So far, only the targeting of EGFR and FRα has been assessed in the clinic, to treat dogs with invasive TCC. A schematic representation of the most promising and advanced targets in the five different cancer types is depicted in Figure 2.
Overview of the molecular targets and targeted therapy drugs for canine and feline cancers discussed in this review
| Molecular target | Homology dog (%)* | Homology cat (%)* | Cancer | Targeted agent | Tested in | Ref. |
|---|---|---|---|---|---|---|
| EGFR | 91.5 | 91.7 | HNSCC | Gefitinib | Feline cells | [44, 45] |
| siRNA | Feline cells | [44] | ||||
| TCC | EGF‐anthrax toxin | Dogs | [49] | |||
| Can225IgG-IRDye700DX | Canine xenograft mouse | [50] | ||||
| OSA | Gefitinib | Canine cells | [54] | |||
| Erlotinib | Canine cells | [55] | ||||
| Erbstatin | Canine cells | [56] | ||||
| MC | Cetuximab | Canine cells | [61] | |||
| Gefitinib | Canine/feline cells | [62] | ||||
| Can225IgG | Canine cells | [63] | ||||
| siRNA | Canine cells | [62] | ||||
| MM | - | - | [67] | |||
| HER2 | 93.7 | 92.9 | TCC | - | - | [70-72] |
| OSA | - | - | [77] | |||
| MC | Trastuzumab | Canine cells | [61] | |||
| AG825 | Canine/feline cells | [62] | ||||
| siRNA | Canine/feline cells | [62] | ||||
| MM | - | - | [67] | |||
| PDGFRα/β | 95.9/90.2 | 95.4/89.8 | OSA | - | - | [91] |
| MM | - | - | [95] | |||
| c-Met | 89.9 | 90.0 | OSA | shRNA | Canine cells | [98] |
| PHA‐665752 | Canine cells | [98] | ||||
| PF2362376 | Canine cells | [101] | ||||
| IGF-1R | 98.2 | 98.1 | OSA | - | - | [103, 104] |
| MM | - | - | [109-111] | |||
| IGF-2R | 85.6 | 85.7 | OSA | - | - | [116] |
| FRα | 80.5 | 75.7 | TCC | EC0905 | Dogs | [118] |
| EC0531 | Dogs | [119] | ||||
| EP2 | 89.0 | 92.1 | TCC | - | - | [72] |
| MCT1/4 | 88.6/87.6 | 88.4/88.0 | HNSCC | MD-1 | Feline xenograft mouse | [124] |
| CD146 | 81.9 | 83.6 | MM | - | - | [127, 128] |
| CXCR4 | 95.8 | 94.9 | MC | - | - | [130-132] |
*The protein sequences of the human, canine and feline molecular targets were found in the Protein NCBI database and aligned with Protein BLAST having human as the reference sequence.
EGFR: epidermal growth factor receptor; HER2: human epidermal growth factor receptor 2; PDGFR-α/β: platelet derived growth factor receptor α/β; c-Met: tyrosine-protein kinase Met; IGF-1/2R: insulin-like growth factor 1/2 receptor; FRα: folate receptor α; EP2: prostaglandin E2 receptor 2; MCT1/4: monocarboxylate transporter 1/4; CD146: cluster of differentiation 146; CXCR4: C-X-C chemokine receptor type 4; HNSCC: head and neck squamous cell carcinoma; TCC: transitional cell carcinoma; OSA: osteosarcoma; MC: mammary carcinoma; MM: malignant melanoma.
Promising molecular targets for the development of anticancer targeted therapies in humans and companion animals. 1. Most advanced and utilized targets with high homology across species for the five most well-characterized cancer types: EGFR and HER2. Other very promising targets: FR, CXCR4, IGFR-1 and PDGFR. 2. Signal deregulation of these membrane proteins, mostly tyrosine kinase receptors, leads to cell proliferation, survival, migration and tumorigenesis. 3. Categories of therapies most often used to interfere with the targets extracellularly or intracellularly. EGFR: epidermal growth factor receptor; HER2: human epidermal growth factor receptor 2; PDGFR: platelet derived growth factor receptor; IGF-1R: insulin-like growth factor 1 receptor; FR: folate receptor; CXCR4: C-X-C chemokine receptor type 4. TKI: tyrosine kinase inhibitor.
As evidenced by this review, EGFR-targeting receives large attention not only in human oncology, but also in veterinary research. Canine and feline EGFR share high homology with the human receptor (92%), making it an interesting target for translational research. In many human cancers, EGFR has proven an invaluable therapeutic target [28]. Multiple EGFR-targeting monoclonal antibodies (e.g. cetuximab and panitumumab) and small molecule inhibitors (e.g. gefitinib and erlotinib) have been approved for the treatment of different human cancers, including HNSCC, colorectal and lung cancer [28, 43]. In this regard, agents targeting EGFR have also been extensively investigated in canine and feline cell lines of OSCC, OSA, TCC and mammary carcinoma, but only the use of EGF‐anthrax toxin has been reported in the clinical setting, for the treatment of canine invasive TCC. This illustrates the large gap between the human and veterinary clinic and points at EGFR targeting as an area where closer collaboration between the human and veterinary clinic could be of great value for companion animals.
The existing distance between human and veterinary oncology is also evident in the treatment of mammary carcinoma. A plethora of anti-HER2 agents are used or are under investigation to treat this human neoplasm; on the contrary, published related research remains at the in vitro phase for canine and feline mammary carcinoma. On a positive note, this opens up avenues for the translation of currently successful and HER2-targeting agents from the human to the veterinary clinic, or even of more affordable generics when available. It is striking to see the lack of research regarding targeting of PDGFRs in canine melanoma, HER2 in canine invasive TCC, CXCR4 in mammary carcinoma and IGF-1R in canine osteosarcoma, all highly expressed molecular targets whose targeting is being evaluated in the human clinic for the respective malignancies. The results of the ongoing human trials, when encouraging, will hopefully spur research in the veterinary setting. In the opposite direction, from the veterinary to the human setting, while targeting of folate receptors has shown great promise in the treatment of dogs with invasive TCC, no reports have been found in human TCC patients and all seems to indicate that also these patients could benefit from the approach.
It is well known that tumor heterogeneity plays a crucial role in the development of drug resistance and patients can clearly benefit from multimodal approaches affecting multiple protumoral pathways. Drawing conclusions from the reviewed data in humans and companion animals, combinations should eventually be considered for the treatment of OSA (including EGFR or PDGFR targeting), HNSCC (with c-Kit inhibition), and mammary carcinoma (using multi-TKIs). As evidenced by this review, the majority of targeted therapies investigated in the veterinary clinic consist of monoclonal antibodies and TKIs. Nowadays, an increasing number of other targeting agents (such as inhibitors of the proteasome and heat shock proteins) are also being assessed, mainly in dogs with hematologic cancers [173]. Advances in the human oncology field and the growing interest in comparative oncology will certainly expand the exploration of more and novel targeted therapies in veterinary patients.
The benefits that can arise from an intricate relationship between human and veterinary clinic are nicely exemplified by the successful story of the drug toceranib phosphate, the first agent approved for anticancer targeted therapy in dogs. The evaluation of toceranib phosphate in dogs with different spontaneous cancers showed the first evidence for the use of an oral TKI as cancer treatment. These findings helped the development of its structural analogue, sunitinib, which is currently approved as a treatment for several human cancers [9]. Interestingly, the analogue intended for human use received approval in 2006, 3 years before toceranib phosphate was approved in the veterinary clinic, which evidences again the reasonably greater interest in human medicine. Overall, this is an example of successful implementation of comparative oncology in the field of targeted therapy, beneficial for both human and veterinary patients. Within this field, other multi-TKI are also starting to show this benefit across species. Dasatinib, already approved in the human clinic, is now being evaluated in veterinary patients [137]. In the opposite direction, masitinib is currently being evaluated in humans, but was first approved to treat companion animals [138]. Another good example of this comparative approach is the case of NHS-IL12, an antibody-cytokine conjugate. This immunotherapy was first evaluated in dogs with melanoma to study its safety, antitumor activity and pharmacological properties, which informed the design of the first-in-human clinical trials [175].
The limited development of targeted therapies in veterinary oncology can be ascribed to several factors: from biological concerns such as the lack of epitope homology between the human and canine/feline target or the hypersensitive reaction to species antibodies, to economic reasons such as a less attractive veterinary oncology market for the industry or the few opportunities for funding [176]. Clinical trials in companion animals allow for data to be collected relatively fast in comparison to human studies. Still, completion of a study may take over a year and carries associated costs, particularly regarding large scale drug production and the clinical services, facilities and instrumentation. Another difficulty is the recruitment of cases for a veterinary trial, which will be dependent on the incidence of the particular cancer and the relatively early diagnosis to allow for treatment benefit. In this respect, a close collaboration with regional veterinary clinics can definitely be of value. One should not forget that treatment-related toxicities are less acceptable in veterinary medicine, a reason why many successful medicines in the human clinic are not being evaluated in veterinary patients. Opportunities may arise here for targeted therapies, which are known to be tumor-selective to some extent, and thus provide a safer setting. To minimize adverse effects, not only is important to verify the presence of the target at the tumor, but also overall target expression and the interactions of the drug with the species target. Although not common, surface expression pattern between species can differ significantly. This led, for instance, to the TGN1412 catastrophe in 2006 [177]. In this respect, theranostic strategies that combine targeted detection before therapy seem a safe approach. Also important is the realization that similar expression pattern does not necessarily translate to identical biological behaviour across species. While most studies in the field rely on immunohistochemistry and PCR techniques to assess expression of a handful of targets, only recently more sophisticated techniques are starting to be introduced that allow for unbiased cross-species analysis, such as wide-genome characterization of the cancer [178] and RNA-sequencing of subpopulations of stroma and neoplastic cells [14, 179]. Such unbiased analysis enables to investigate similarities between species and to identify new therapeutic targets.
Altogether, when relevant and feasible, breaking the current linearity of drug development programs (mainly from rodents to humans) by incorporating veterinary patients can guide the successful implementation of therapeutics in humans, or aid in identifying failure earlier.
Conclusion
The use of comparative oncology for translational research of targeted therapies is of benefit for both human and veterinary patients. While improving the translational success rate of these therapies from preclinical to clinical studies, dogs and cats get access to otherwise inaccessible novel therapies. Equally important, targeting therapeutic agents approved for human use can be a potential treatment for dogs and cats with tumors expressing the molecular target in question. Clearly, researchers in the field of veterinary and human oncology could help each other and, more importantly, their patients by working closely together.
Abbreviations
BRAF: B-Raf proto-oncogene; CMC: canine mammary carcinoma; COX2: cyclooxygenase 2; CXCR4: C-X-C chemokine receptor type 4; EGFR: epidermal growth factor receptor; EP2: prostaglandin E2 receptor 2; FMC: feline mammary carcinoma; FGFR: fibroblast growth factor receptor; FR: folate receptor; HER2: human epidermal growth factor receptor 2; HNSCC: head and neck squamous cell carcinoma; IGF: insulin-like growth factor; IGF-1/2R: insulin-like growth factor 1/2 receptor; MCT: monocarboxylate transporter; MEK: mitogen-activated protein kinase; NSAID: non-steroidal anti-inflammatory drugs; OSA: osteosarcoma; OSCC: oral squamous cell carcinoma; PDGFR-α/β: platelet derived growth factor receptor α/β; TCC: transitional cell carcinoma; TKI: tyrosine kinase inhibitor; VEGF: vascular endothelial growth factor; VEGFR: vascular endothelial growth factor receptor.
Acknowledgements
This work was supported by the European Research Council (ERC) under the European Union's Horizon 2020 Research and Innovation Program (Grant agreement No. 677582).
Competing Interests
The authors have declared that no competing interest exists.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394-424
2. Miller KD, Nogueira L, Mariotto AB, Rowland JH, Yabroff KR, Alfano CM. et al. Cancer treatment and survivorship statistics, 2019. CA Cancer J Clin. 2019;69:363-385
3. Wolchok J. Putting the Immunologic Brakes on Cancer. Cell. 2018;175:1452-1454
4. Lee YT, Tan YJ, Oon CE. Molecular targeted therapy: Treating cancer with specificity. Eur J Pharmacol. 2018;834:188-196
5. Heem Wong C, Wei Siah K, Lo AW. Estimation of clinical trial success rates and related parameters. Biostatistics. 2019;20:273-286
6. Mak IWY, Evaniew N, Ghert M. Lost in translation: animal models and clinical trials in cancer treatment. Am J Transl Res. 2014;6:114-118
7. Garden OA, Volk SW, Mason NJ, Perry JA. Companion animals in comparative oncology: One Medicine in action. Vet J. 2018;240:6-13
8. Riccardo F, Aurisicchio L, Impellizeri JA, Cavallo F. The importance of comparative oncology in translational medicine. Cancer Immunol Immunother. 2015;64:137-148
9. Regan D, Garcia K, Thamm D. Clinical, pathological, and ethical considerations for the conduct of clinical trials in dogs with naturally occurring cancer: a comparative approach to accelerate translational drug development. ILAR J. 2018;59:99-110
10. Cannon CM. Cats, Cancer and Comparative Oncology. Vet Sci. 2015;2:111-126
11. Lindblad-Toh K, Wade CM, Mikkelsen TS, Karlsson EK, Jaffe DB, Kamal M. et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature. 2005;438:803-819
12. Montague MJ, Li G, Gandolfi B, Khan R, Aken BL, Searle SMJ. et al. Comparative analysis of the domestic cat genome reveals genetic signatures underlying feline biology and domestication. Proc Natl Acad Sci. 2014;111:17230 LP - 17235
13. Supsavhad W, Dirksen WP, Martin CK, Rosol TJ. Animal models of head and neck squamous cell carcinoma. Vet J. 2016;210:7-16
14. Guscetti F, Nassiri S, Beebe E, Brandao IR, Graf R, Markkanen E. Molecular homology between canine spontaneous oral squamous cell carcinomas and human head-and- neck squamous cell carcinomas reveals disease drivers and therapeutic vulnerabilities. Neoplasia. 2020;22:778-788
15. Wypij JM. A naturally occurring feline model of head and neck squamous cell carcinoma. Patholog Res Int. 2013;2013:502197
16. Argyle DJ. The Webinar Vet. https://www.thewebinarvet.com/pages/cancers-head-neck/.
17. Knapp DW, Dhawan D, Ramos-Vara JA, Ratliff TL, Cresswell GM, Utturkar S. et al. Naturally-Occurring Invasive Urothelial Carcinoma in Dogs, a Unique Model to Drive Advances in Managing Muscle Invasive Bladder Cancer in Humans. Front Oncol. 2020;9:1493
18. Gray M, Meehan J, Martínez-Pérez C, Kay C, Turnbull AK, Morrison LR. et al. Naturally-Occurring Canine Mammary Tumors as a Translational Model for Human Breast Cancer. Front Oncol. 2020;10:1-17
19. Hassan BB, Elshafae SM, Supsavhad W, Simmons JK, Dirksen WP, Sokkar SM. et al. Feline mammary cancer: novel nude mouse model and molecular characterization of invasion and metastasis genes. Vet Pathol. 2016;54:32-43
20. Mirabello L, Troisi RJ, Savage SA. International osteosarcoma incidence patterns in children and adolescents, middle ages, and elderly persons. Int J Cancer. 2009;125:229-234
21. Simpson S, Dunning MD, de Brot S, Grau-Roma L, Mongan NP, Rutland CS. Comparative review of human and canine osteosarcoma: morphology, epidemiology, prognosis, treatment and genetics. Acta Vet Scand. 2017 59
22. Gustafson DL, Duval DL, Regan DP, Thamm DH. Canine sarcomas as a surrogate for the human disease. Pharmacol Ther. 2018;188:80-96
23. Skin Cancer Foundation. https://www.skincancer.org/skin-cancer-information/skin-cancer-facts/
24. Prouteau A, André C. Canine Melanomas as Models for Human Melanomas: Clinical, Histological, and Genetic Comparison. Genes (Basel). 2019 10
25. Gillard M, Cadieu E, Brito C De, Abadie J, Vergier B, Devauchelle P. et al. Naturally occurring melanomas in dogs as models for non-UV pathways of human melanomas. Pigment Cell Melanoma Res. 2014;27:90-102
26. Pang L, Argyle D. Veterinary oncology: biology, big data and precision medicine. Vet J. 2016;213:38-45
27. FETCH a Cure. https://fetchacure.org/resource-library/facts
28. Jue Xu M, Johnson DE, Grandis JR. EGFR-Targeted Therapies in the Post-Genomic Era. Cancer Metastasis Rev. 2017;36:463-473
29. Ernberg IT. Oncogenes and tumor growth factors in breast cancer. A minireview. Acta Oncol (Madr). 1990;29:331-334
30. Pietras K, Sjöblom T, Rubin K, Heldin C, Östman A. PDGF receptors as cancer drug targets. Cancer Cell. 2003;3:439-443
31. Zhang Y, Xia M, Jin K, Wang S, Wei H, Fan C. et al. Function of the c-Met receptor tyrosine kinase in carcinogenesis and associated therapeutic opportunities. Mol Cancer. 2018 17
32. Osher E, Macaulay VM. Therapeutic targeting of the IGF axis. Cells. 2019;8:895
33. Martin-Kleiner I, Gall Troselj K. Mannose-6-phosphate/insulin-like growth factor 2 receptor (M6P/IGF2R) in carcinogenesis. Cancer Lett. 2010;289:11-22
34. Scaranti M, Cojocaru E, Banerjee S, Banerji U. Exploiting the folate receptor α in oncology. Nat Rev Clin Oncol. 2020
35. Sun X, Li Q. Prostaglandin EP2 receptor: Novel therapeutic target for human cancers (Review). Int J Mol Med. 2018;42:1203-1214
36. Halestrap AP. The SLC16 gene family - Structure, role and regulation in health and disease. Mol Aspects Med. 2013;34:337-349
37. Lei X, Guan C-W, Song Y, Wang H. The multifaceted role of CD146/MCAM in the promotion of melanoma progression. Cancer Cell Int. 2015;15:3
38. Zhou W, Guo S, Liu M, Burow ME, Wang G. Targeting CXCL12/CXCR4 axis in tumor immunotherapy. Curr Med Chem. 2019;26:3026-3041
39. Looper JS, Malarkey DE, Ruslander D, Proulx D, Thrall DE. Epidermal growth factor receptor expression in feline oral squamous cell carcinomas. Vet Comp Oncol. 2006;4:33-41
40. Bergkvist GT, Argyle DJ, Morrison L, MacIntyre N, Hayes A, Yool DA. Expression of epidermal growth factor receptor (EGFR) and Ki67 in feline oral squamous cell carcinomas (FOSCC). Vet Comp Oncol. 2011;9:106-117
41. Bossi P, Resteghini C, Paielli N, Licitra L, Pilotti S, Perrone F. Prognostic and predictive value of EGFR in head and neck squamous cell carcinoma. Oncotarget. 2016;7:74362-74379
42. Kozakiewicz P, Grzybowska-Szatkowska L. Application of molecular targeted therapies in the treatment of head and neck squamous cell carcinoma. Oncol Lett. 2018;15:7497-7505
43. Cohen MH, Chen H, Shord S, Fuchs C, He K, Zhao H. et al. Approval summary: Cetuximab in combination with cisplatin or carboplatin and 5-fluorouracil for the first-line treatment of patients with recurrent locoregional or metastatic squamous cell head and neck cancer. Oncologist. 2013;18:460-466
44. Bergkvist GT, Argyle DJ, Pang LY, Muirhead R, Yool DA. Studies on the inhibition of feline EGFR in squamous cell carcinoma: Enhancement of radiosensitivity and rescue of resistance to small molecule inhibitors. Cancer Biol Ther. 2011;11:927-937
45. Pang LY, Bergkvist GT, Cervantes-Arias A, Yool DA, Muirhead R, Argyle DJ. Identification of tumour initiating cells in feline head and neck squamous cell carcinoma and evidence for gefitinib induced epithelial to mesenchymal transition. Vet J. 2012;193:46-52
46. Hanazono K, Fukumoto S, Kawamura Y, Endo Y, Kadosawa T, Iwano H. et al. Epidermal growth factor receptor expression in canine transitional cell carcinoma. J Vet Med Sci. 2015;77:1-6
47. Wright C, Mellon K, Johnston P, Lane DP, Harris AL, Horne CH. et al. Expression of mutant p53, c-erbB-2 and the epidermal growth factor receptor in transitional cell carcinoma of the human urinary bladder. Br J Cancer. 1991;63:967-970
48. Goodspeed A, Jean A, Theodorescu D, Costello JC. A gene expression signature predicts bladder cancer cell line sensitivity to EGFR inhibition. Bl cancer (Amsterdam, Netherlands). 2018;4:269-282
49. Jack S, Madhivanan K, Ramadesikan S, Subramanian S, Edwards II DF, Elzey BD. et al. A novel, safe, fast and efficient treatment for Her2-positive and negative bladder cancer utilizing an EGF-anthrax toxin chimera. Int J Cancer. 2020;146:449-460
50. Nagaya T, Okuyama S, Ogata F, Maruoka Y, Knapp DW, Karagiannis SN. et al. Near infrared photoimmunotherapy targeting bladder cancer with a canine anti-epidermal growth factor receptor (EGFR) antibody. Oncotarget. 2018;9:19026-19038
51. Wen YH, Koeppen H, Garcia R, Chiriboga L, Tarlow BD, Peters BA. et al. Epidermal growth factor receptor in osteosarcoma: expression and mutational analysis. Hum Pathol. 2007;38:1184-1191
52. Wang W, Zhao H, Yao T, Gong H. Advanced development of ErbB family-targeted therapies in osteosarcoma treatment. Invest New Drugs. 2019;37:175-183
53. Selvarajah GT, Verheije MH, Kik M, Slob A, Rottier PJM, Mol JA. et al. Expression of epidermal growth factor receptor in canine osteosarcoma: Association with clinicopathological parameters and prognosis. Vet J. 2012;193:412-419
54. McCleese JK, Bear MD, Kulp SK, Mazcko C, Khanna C, London CA. Met interacts with EGFR and Ron in canine osteosarcoma. Vet Comp Oncol. 2013;11:124-139
55. Mantovani FB, Morrison JA, Mutsaers AJ. Effects of epidermal growth factor receptor kinase inhibition on radiation response in canine osteosarcoma cells. BMC Vet Res. 2016;12:82
56. Mauchle U, Selvarajah GT, Mol JA, Kirpensteijn J, Verheije MH. Identification of anti-proliferative kinase inhibitors as potential therapeutic agents to treat canine osteosarcoma. Vet J. 2015;205:281-287
57. Rimawi MF, Shetty PB, Weiss HL, Schiff R, Osborne CK, Chamness GC. et al. Epidermal growth factor receptor expression in breast cancer association with biologic phenotype and clinical outcomes. Cancer. 2010;116:1234-1242
58. Maennling AE, Tur MK, Niebert M, Klockenbring T, Zeppernick F, Gattenlöhner S. et al. Molecular targeting therapy against EGFR family in breast cancer: progress and future potentials. Cancers (Basel). 2019;11:1826
59. Gama A, Gärtner F, Alves A, Schmitt F. Immunohistochemical expression of Epidermal Growth Factor Receptor (EGFR) in canine mammary tissues. Res Vet Sci. 2009;87:432-437
60. Hassan BB, Elshafae SM, Supsavhad W, Simmons JK, Dirksen WP, Sokkar SM. et al. Feline mammary cancer. Vet Pathol. 2017;54:32-43
61. Singer J, Weichselbaumer M, Stockner T, Mechtcheriakova D, Sobanov Y, Bajna E. et al. Comparative oncology: ErbB-1 and ErbB-2 homologues in canine cancer are susceptible to cetuximab and trastuzumab targeting. Mol Immunol. 2012;50:200-209
62. Gray ME, Lee S, McDowell AL, Erskine M, Loh QTM, Grice O. et al. Dual targeting of EGFR and ERBB2 pathways produces a synergistic effect on cancer cell proliferation and migration in vitro. Vet Comp Oncol. 2017;15:890-909
63. Singer J, Fazekas J, Wang W, Weichselbaumer M, Matz M, Mader A. et al. Generation of a canine anti-EGFR (ErbB-1) antibody for passive immunotherapy in dog cancer patients. Mol Cancer Ther. 2014;13:1777-1790
64. Boone B, Jacobs K, Ferdinande L, Taildeman J, Lambert J, Peeters M. et al. EGFR in melanoma: clinical significance and potential therapeutic target. J Cutan Pathol. 2011;38:492-502
65. Patel SP, Kim KB, Papadopoulos NE, Hwu W-J, Hwu P, Prieto VG. et al. A phase II study of gefitinib in patients with metastatic melanoma. Melanoma Res. 2011;21:357-363
66. Mudigonda T V, Wyman K, Spigel DR, Dahlman KB, Greco FA, Puzanov I. et al. A phase II trial of erlotinib and bevacizumab for patients with metastatic melanoma. Pigment Cell Melanoma Res. 2016;29:101-103
67. Veloso ES, Gonçalves INN, Silveira TL, Oliveira FS, Vieira DS, Cassali GD. et al. Diverse roles of epidermal growth factors receptors in oral and cutaneous canine melanomas. BMC Vet Res. 2020;16:24
68. Rahman MM, Lai Y-C, Husna AA, Chen H-W, Tanaka Y, Kawaguchi H. et al. Transcriptome analysis of dog oral melanoma and its oncogenic analogy with human melanoma. Oncol Rep. 2020;43:16-30
69. Coogan CL, Estrada CR, Kapur S, Bloom KJ. HER-2/neu protein overexpression and gene amplification in human transitional cell carcinoma of the bladder. Urology. 2004;63:786-790
70. Millanta F, Impellizeri J, McSherry L, Rocchigiani G, Aurisicchio L, Lubas G. Overexpression of HER-2 via immunohistochemistry in canine urinary bladder transitional cell carcinoma - A marker of malignancy and possible therapeutic target. Vet Comp Oncol. 2018;16:297-300
71. Tsuboi M, Sakai K, Maeda S, Chambers JK, Yonezawa T, Matsuki N. et al. Assessment of HER2 expression in canine urothelial carcinoma of the urinary bladder. Vet Pathol. 2019;56:369-376
72. Maeda S, Tomiyasu H, Tsuboi M, Inoue A, Ishihara G, Uchikai T. et al. Comprehensive gene expression analysis of canine invasive urothelial bladder carcinoma by RNA-Seq. BMC Cancer. 2018;18:472
73. Koshkin VS, O'Donnell P, Yu EY, Grivas P. Systematic review: Targeting HER2 in bladder cancer. Bl Cancer. 2019;5:1-12
74. Tabak SA, Khalifa SE, Fathy Y. HER-2 immunohistochemical expression in bone sarcomas: a new hope for osteosarcoma patients. Open access Maced J Med Sci. 2018;6:1555-1560
75. Thomas DG, Giordano TJ, Sanders D, Biermann JS, Baker L. Absence of HER2/neu gene expression in osteosarcoma and skeletal Ewings sarcoma. Clin Cancer Res. 2002;8:788-793
76. Kilpatrick SE, Geisinger KR, King TS, Sciarrotta J, Ward WG, Gold SH. et al. Clinicopathologic analysis of HER-2/neu immunoexpression among various histologic subtypes and grades of osteosarcoma. Mod Pathol. 2001;14:1277-1283
77. Flint AF, U'Ren L, Legare ME, Withrow SJ, Dernell W, Hanneman WH. Overexpression of the erbb-2 proto-oncogene in canine osteosarcoma cell lines and tumors. Vet Pathol. 2004;41:291-296
78. Ebb D, Meyers P, Grier H, Bernstein M, Gorlick R, Lipshultz SE. et al. Phase II trial of trastuzumab in combination with cytotoxic chemotherapy for treatment of metastatic osteosarcoma with human epidermal growth factor receptor 2 overexpression: a report from the children's oncology group. J Clin Oncol. 2012;30:2545-2551
79. Ahmed N, Brawley VS, Hegde M, Robertson C, Ghazi A, Gerken C. et al. Human epidermal growth factor receptor 2 (HER2) -specific chimeric antigen receptor-modified T Cells for the immunotherapy of HER2-positive sarcoma. J Clin Oncol. 2015;33:1688-1696
80. Mason NJ, Gnanandarajah JS, Engiles JB, Gray F, Laughlin D, Gaurnier-Hausser A. et al. Immunotherapy with a HER2-Targeting Listeria Induces HER2-Specific Immunity and Demonstrates Potential Therapeutic Effects in a Phase I Trial in Canine Osteosarcoma. Clin Cancer Res. 2016;22:4380 LP - 4390
81. Ménard S, Tagliabue E, Campiglio M, Pupa SM. Role of HER2 gene overexpression in breast carcinoma. J Cell Physiol. 2000;182:150-162
82. Marques C, Correia J, Ferreira F. HER2-positive feline mammary carcinoma. Aging (Albany NY). 2016;8:1574-1575
83. De Maria R, Olivero M, Iussich S, Nakaichi M, Murata T, Biolatti B. et al. Spontaneous feline mammary carcinoma is a model of HER2 overexpressing poor prognosis human breast cancer. Cancer Res. 2005;65:907 LP - 912
84. Ressel L, Puleio R, Loria GR, Vannozzi I, Millanta F, Caracappa S. et al. HER-2 expression in canine morphologically normal, hyperplastic and neoplastic mammary tissues and its correlation with the clinical outcome. Res Vet Sci. 2013;94:299-305
85. Hsu W-L, Huang H-M, Liao J-W, Wong M-L, Chang S-C. Increased survival in dogs with malignant mammary tumours overexpressing HER-2 protein and detection of a silent single nucleotide polymorphism in the canine HER-2 gene. Vet J. 2009;180:116-123
86. Kluger HM, DiVito K, Berger AJ, Halaban R, Ariyan S, Camp RL. et al. Her2/neu is not a commonly expressed therapeutic target in melanoma - a large cohort tissue microarray study. Melanoma Res. 2004 14
87. Bodey B, Bodey JB, Gröger AM, Luck J V, Siegel SE, Taylor CR. et al. Clinical and prognostic significance of the expression of the c-erbB-2 and c-erbB-3 oncoproteins in primary and metastatic malignant melanomas and breast carcinomas. Anticancer Res. 1997;17:1319-1330
88. Ma J, Han H, Liu D, Li W, Feng H, Xue X. et al. HER2 as a promising target for cytotoxicity T cells in human melanoma therapy. PLoS One. 2013;8:e73261-e73261
89. Kubo T, Piperdi S, Rosenblum J, Antonescu CR, Chen W, Kim H-S. et al. Platelet-derived growth factor receptor as a prognostic marker and a therapeutic target for imatinib mesylate therapy in osteosarcoma. Cancer. 2008;112:2119-2129
90. Xu J, Xie L, Guo W. PDGF/PDGFR effects in osteosarcoma and the “add-on” strategy. Clin Sarcoma Res. 2018;8:15
91. Maniscalco L, Iussich S, Morello E, Martano M, Biolatti B, Riondato F. et al. PDGFs and PDGFRs in canine osteosarcoma: New targets for innovative therapeutic strategies in comparative oncology. Vet J. 2013;195:41-47
92. D'Arcangelo D, Facchiano F, Nassa G, Stancato A, Antonini A, Rossi S. et al. PDGFR-alpha inhibits melanoma growth via CXCL10/IP-10: a multi-omics approach. Oncotarget. 2016;7:77257-77275
93. Barnhill RL, Xiao M, Graves D, Antoniades HN. Expression of platelet-derived growth factor (PDGF)-A, PDGF-B and the PDGF-alpha receptor, but not the PDGF-beta receptor, in human malignant melanoma in vivo. Br J Dermatol. 1996;135:898-904
94. Papadopoulos N, Lennartsson J. The PDGF/PDGFR pathway as a drug target. Mol Aspects Med. 2018;62:75-88
95. Iussich S, Maniscalco L, Di Sciuva A, Iotti B, Morello E, Martano M. et al. PDGFRs expression in dogs affected by malignant oral melanomas: correlation with prognosis. Vet Comp Oncol. 2017;15:462-469
96. Hassan SE, Bekarev M, Kim MY, Lin J, Piperdi S, Gorlick R. et al. Cell surface receptor expression patterns in osteosarcoma. Cancer. 2012;118:740-749
97. Scotlandi K, Baldini N, Oliviero M, Di Renzo MF, Martano M, Serra M. et al. Expression of Met/hepatocyte growth factor receptor gene and malignant behavior of musculoskeletal tumors. Am J Pathol. 1996;149:1209-1219
98. De Maria R, Miretti S, Iussich S, Olivero M, Morello E, Bertotti A. et al. met oncogene activation qualifies spontaneous canine osteosarcoma as a suitable pre-clinical model of human osteosarcoma. J Pathol. 2009;218:399-408
99. Fioramonti M, Fausti V, Pantano F, Iuliani M, Ribelli G, Lotti F. et al. Cabozantinib affects osteosarcoma growth through a direct effect on tumor cells and modifications in bone microenvironment. Sci Rep. 2018;8:4177
100. Sampson ER, Martin BA, Morris AE, Xie C, Schwarz EM, O'Keefe RJ. et al. The orally bioavailable met inhibitor PF-2341066 inhibits osteosarcoma growth and osteolysis/matrix production in a xenograft model. J Bone Miner Res. 2011;26:1283-1294
101. Liao AT, McCleese J, Kamerling S, Christensen J, London CA. A novel small molecule Met inhibitor, PF2362376, exhibits biological activity against osteosarcoma. Vet Comp Oncol. 2007;5:177-196
102. Wang Y-H, Han X-D, Qiu Y, Xiong J, Yu Y, Wang B. et al. Increased expression of insulin-like growth factor-1 receptor is correlated with tumor metastasis and prognosis in patients with osteosarcoma. J Surg Oncol. 2012;105:235-243
103. MacEwen EG, Pastor J, Kutzke J, Tsan R, Kurzman ID, Thamm DH. et al. IGF-1 receptor contributes to the malignant phenotype in human and canine osteosarcoma. J Cell Biochem. 2004;92:77-91
104. Maniscalco L, Iussich S, Morello E, Martano M, Gattino F, Miretti S. et al. Increased expression of insulin-like growth factor-1 receptor is correlated with worse survival in canine appendicular osteosarcoma. Vet J. 2015;205:272-280
105. Pappo AS, Vassal G, Crowley JJ, Bolejack V, Hogendoorn PCW, Chugh R. et al. A phase 2 trial of R1507, a monoclonal antibody to the insulin-like growth factor-1 receptor (IGF-1R), in patients with recurrent or refractory rhabdomyosarcoma, osteosarcoma, synovial sarcoma, and other soft tissue sarcomas: results of a Sarcoma Alliance. Cancer. 2014;120:2448-2456
106. Li Y, Liu Q, He H, Luo W. The possible role of insulin-like growth factor-1 in osteosarcoma. Curr Probl Cancer. 2019;43:228-235
107. Yerushalmi R, Gelmon KA, Leung S, Gao D, Cheang M, Pollak M. et al. Insulin-like growth factor receptor (IGF-1R) in breast cancer subtypes. Breast Cancer Res Treat. 2012;132:131-142
108. Ekyalongo RC, Yee D. Revisiting the IGF-1R as a breast cancer target. NPJ Precis Oncol. 2017;1:14
109. Jaillardon L, Abadie J, Godard T, Campone M, Loussouarn D, Siliart B. et al. The dog as a naturally-occurring model for insulin-like growth factor type 1 receptor-overexpressing breast cancer: an observational cohort study. BMC Cancer. 2015;15:664
110. Dolka I, Motyl T, Malicka E, Sapierzyński R, Fabisiak M. Relationship between receptors for insulin-like growth factor-I, steroid hormones and apoptosis-associated proteins in canine mammary tumors. Pol J Vet Sci. 2011;14:245-251
111. Klopfleisch R, Hvid H, Klose P, Da Costa A, Gruber AD. Insulin receptor is expressed in normal canine mammary gland and benign adenomas but decreased in metastatic canine mammary carcinomas similar to human breast cancer. Vet Comp Oncol. 2010;8:293-301
112. Kanter-Lewensohn L, Dricu A, Girnita L, Wejde J, Larsson O. Expression of insulin-like growth factor-1 receptor (IGF-1R) and p27Kip1 in melanocyte tumors: a potential regulatory role of IGF-1 pathway in distribution of p27Kip1 between different cyclins. Growth Factors. 2000;17:193-202
113. Capoluongo E. Insulin-like growth factor system and sporadic malignant melanoma. Am J Pathol. 2011;178:26-31
114. Thamm DH, Huelsmeyer MK, Mitzey AM, Qurollo B, Rose BJ, Kurzman ID. RT-PCR-based tyrosine kinase display profiling of canine melanoma: IGF-1 receptor as a potential therapeutic target. Melanoma Res. 2010 20
115. Simpson A, Petnga W, Macaulay VM, Weyer-Czernilofsky U, Bogenrieder T. Insulin-like growth factor (IGF) pathway targeting in cancer: Role of the IGF axis and opportunities for future combination studies. Target Oncol. 2017;12:571-597
116. Karkare S, Allen KJH, Jiao R, Malo ME, Dawicki W, Helal M. et al. Detection and targeting insulin growth factor receptor type 2 (IGF2R) in osteosarcoma PDX in mouse models and in canine osteosarcoma tumors. Sci Rep. 2019;9:11476
117. Geller DS, Morris J, Revskaya E, Kahn M, Zhang W, Piperdi S. et al. Targeted therapy of osteosarcoma with radiolabeled monoclonal antibody to an insulin-like growth factor-2 receptor (IGF2R). Nucl Med Biol. 2016;43:812-817
118. Dhawan D, Ramos-Vara JA, Naughton JF, Cheng L, Low PS, Rothenbuhler R. et al. Targeting folate receptors to treat invasive urinary bladder cancer. Cancer Res. 2013;73:875 LP - 884
119. Szigetvari NM, Dhawan D, Ramos-Vara JA, Leamon CP, Klein PJ, Ruple AA. et al. Phase I/II clinical trial of the targeted chemotherapeutic drug, folate-tubulysin, in dogs with naturally-occurring invasive urothelial carcinoma. Oncotarget. 2018;9:37042-37053
120. Kashiwagi E, Inoue S, Mizushima T, Chen J, Ide H, Kawahara T. et al. Prostaglandin receptors induce urothelial tumourigenesis as well as bladder cancer progression and cisplatin resistance presumably via modulating PTEN expression. Br J Cancer. 2018;118:213-223
121. von der Emde L, Goltz D, Latz S, Müller SC, Kristiansen G, Ellinger J. et al. Prostaglandin receptors EP1-4 as a potential marker for clinical outcome in urothelial bladder cancer. Am J Cancer Res. 2014;4:952-962
122. Simões-Sousa S, Granja S, Pinheiro C, Fernandes D, Longatto-Filho A, Laus AC. et al. Prognostic significance of monocarboxylate transporter expression in oral cavity tumors. Cell Cycle. 2016;15:1865-1873
123. Curry JM, Tuluc M, Whitaker-Menezes D, Ames JA, Anantharaman A, Butera A. et al. Cancer metabolism, stemness and tumor recurrence: MCT1 and MCT4 are functional biomarkers of metabolic symbiosis in head and neck cancer. Cell Cycle. 2013;12:1371-1384
124. Khammanivong A, Saha J, Spartz AK, Sorenson BS, Bush AG, Korpela DM. et al. A novel MCT1 and MCT4 dual inhibitor reduces mitochondrial metabolism and inhibits tumour growth of feline oral squamous cell carcinoma. Vet Comp Oncol. 2019 18
125. Pearl RA, Pacifico MD, Richman PI, Wilson GD, Grover R. Stratification of patients by melanoma cell adhesion molecule (MCAM) expression on the basis of risk: implications for sentinel lymph node biopsy. J Plast Reconstr Aesthetic Surg. 2008;61:265-271
126. Wei W, Jiang D, Ehlerding EB, Barnhart TE, Yang Y, Engle JW. et al. CD146-targeted multimodal image-guided photoimmunotherapy of melanoma. Adv Sci. 2019;6:1801237
127. Abou Asa S. Immunohistochemical expression of MCAM/CD146 in canine melanoma. J Comp Pathol. 2017;157:27-33
128. Schmid F, Brodesser D, Reifinger M, Forte S, Semp P, Eberspächer-Schweda MC. et al. Canine oral primary melanoma cells exhibit shift to mesenchymal phenotype and phagocytic behaviour. Vet Comp Oncol. 2019;17:211-220
129. Pernas S, Martin M, Kaufman PA, Gil-Martin M, Gomez Pardo P, Lopez-Tarruella S. et al. Balixafortide plus eribulin in HER2-negative metastatic breast cancer: a phase 1, single-arm, dose-escalation trial. Lancet Oncol. 2018;19:812-824
130. Marques CS, Santos AR, Gameiro A, Correia J, Ferreira F. CXCR4 and its ligand CXCL12 display opposite expression profiles in feline mammary metastatic disease, with the exception of HER2-overexpressing tumors. BMC Cancer. 2018;18:741
131. Ferrari A, Petterino C, Ratto A, Campanella C, Wurth R, Thellung S. et al. CXCR4 expression in feline mammary carcinoma cells: evidence of a proliferative role for the SDF-1/CXCR4 axis. BMC Vet Res. 2012;8:27
132. Hussain S, Saxena S, Shrivastava S, Mohanty AK, Kumar S, Singh RJ. et al. Gene expression profiling of spontaneously occurring canine mammary tumours: Insight into gene networks and pathways linked to cancer pathogenesis. PLoS One. 2018;13:e0208656-e0208656
133. Gschwind A, Fischer OM, Ullrich A. The discovery of receptor tyrosine kinases: targets for cancer therapy. Nat Rev Cancer. 2004;4:361-370
134. Wiles V, Hohenhaus A, Lamb K, Zaidi B, Camps-Palau M, Leibman N. Retrospective evaluation of toceranib phosphate (Palladia) in cats with oral squamous cell carcinoma. J Feline Med Surg. 2016;19:185-193
135. London C, Mathie T, Stingle N, Clifford C, Haney S, Klein MK. et al. Preliminary evidence for biologic activity of toceranib phosphate (Palladia(®)) in solid tumours. Vet Comp Oncol. 2012;10:194-205
136. Papaetis GS, Syrigos KN. Sunitinib. BioDrugs. 2009;23:377-389
137. Montero JC, Seoane S, Ocaña A, Pandiella A. Inhibition of Src family kinases and receptor tyrosine kinases by dasatinib: possible combinations in solid tumors. Clin Cancer Res. 2011;17:5546 LP - 5552
138. Dubreuil P, Letard S, Ciufolini M, Gros L, Humbert M, Castéran N. et al. Masitinib (AB1010), a potent and selective tyrosine kinase inhibitor targeting KIT. PLoS One. 2009;4:e7258-e7258
139. Rathore K, Alexander M, Cekanova M. Piroxicam inhibits Masitinib-induced cyclooxygenase 2 expression in oral squamous cell carcinoma cells in vitro. Transl Res. 2014;164:158-168
140. Gustafson TL, Biller B. Use of toceranib phosphate in the treatment of canine bladder tumors: 37 cases. J Am Anim Hosp Assoc. 2019;55:243-248
141. Davis LE, Hofmann NE, Li G, Huang ET, Loriaux MM, Bracha S. et al. A case study of personalized therapy for osteosarcoma. Pediatr Blood Cancer. 2013;60:1313-1319
142. Marley K, Gullaba J, Seguin B, Gelberg HB, Helfand SC. Dasatinib modulates invasive and migratory properties of canine osteosarcoma and has therapeutic potential in affected dogs. Transl Oncol. 2015;8:231-238
143. Rossi F, Sabattini S, Vascellari M, Marconato L. The impact of toceranib, piroxicam and thalidomide with or without hypofractionated radiation therapy on clinical outcome in dogs with inflammatory mammary carcinoma. Vet Comp Oncol. 2018;16:497-504
144. London CA, Hannah AL, Zadovoskaya R, Chien MB, Kollias-Baker C, Rosenberg M. et al. Phase I dose-escalating study of SU11654, a small molecule receptor tyrosine kinase inhibitor, in dogs with spontaneous malignancies. Clin Cancer Res. 2003;9:2755-2768
145. Wouda RM, Hocker SE, Higginbotham ML. Safety evaluation of combination carboplatin and toceranib phosphate (Palladia) in tumour-bearing dogs: A phase I dose finding study. Vet Comp Oncol. 2018;16:E52-E60
146. Choong NW, Kozloff M, Taber D, Hu HS, Wade J, Ivy P. et al. Phase II study of sunitinib malate in head and neck squamous cell carcinoma. Invest New Drugs. 2010;28:677-683
147. Machiels J-PH, Henry S, Zanetta S, Kaminsky M-C, Michoux N, Rommel D. et al. Phase II study of sunitinib in recurrent or metastatic squamous cell carcinoma of the head and neck: GORTEC 2006-01. J Clin Oncol. 2009;28:21-28
148. Zehnder A, Graham J, Antonissen G. Update on cancer treatment in exotics. Vet Clin North Am Exot Anim Pract. 2018;21:465-509
149. Ongkeko WM, Altuna X, Weisman RA, Wang-Rodriguez J. Expression of protein tyrosine kinases in head and neck squamous cell carcinomas. Am J Clin Pathol. 2005;124:71-76
150. Zahoor H, Mir MC, Barata PC, Stephenson AJ, Campbell SC, Fergany A. et al. Phase II trial of continuous treatment with sunitinib in patients with high-risk (BCG-refractory) non-muscle invasive bladder cancer. Invest New Drugs. 2019;37:1231-1238
151. Grivas PD, Daignault S, Tagawa ST, Nanus DM, Stadler WM, Dreicer R. et al. Double-blind, randomized, phase 2 trial of maintenance sunitinib versus placebo after response to chemotherapy in patients with advanced urothelial carcinoma. Cancer. 2014;120:692-701
152. Bellmunt J, González-Larriba JL, Prior C, Maroto P, Carles J, Castellano D. et al. Phase II study of sunitinib as first-line treatment of urothelial cancer patients ineligible to receive cisplatin-based chemotherapy: baseline interleukin-8 and tumor contrast enhancement as potential predictive factors of activity. Ann Oncol. 2011;22:2646-2653
153. Walters L, Martin O, Price J, Sula MM. Expression of receptor tyrosine kinase targets PDGFR-β, VEGFR2 and KIT in canine transitional cell carcinoma. Vet Comp Oncol. 2018;16:E117-E122
154. Kumar RMR, Arlt MJ, Kuzmanov A, Born W, Fuchs B. Sunitinib malate (SU-11248) reduces tumour burden and lung metastasis in an intratibial human xenograft osteosarcoma mouse model. Am J Cancer Res. 2015;5:2156-2168
155. Gieger TL, Nettifee-Osborne J, Hallman B, Johannes C, Clarke D, Nolan MW. et al. The impact of carboplatin and toceranib phosphate on serum vascular endothelial growth factor (VEGF) and metalloproteinase-9 (MMP-9) levels and survival in canine osteosarcoma. Can J Vet Res. 2017;81:199-205
156. Kim C, Matsuyama A, Mutsaers AJ, Woods JP. Retrospective evaluation of toceranib (Palladia) treatment for canine metastatic appendicular osteosarcoma. Can Vet J = La Rev Vet Can. 2017;58:1059-1064
157. Laver T, London CA, Vail DM, Biller BJ, Coy J, Thamm DH. Prospective evaluation of toceranib phosphate in metastatic canine osteosarcoma. Vet Comp Oncol. 2018;16:E23-E29
158. London CA, Gardner HL, Mathie T, Stingle N, Portela R, Pennell ML. et al. Impact of toceranib/piroxicam/cyclophosphamide maintenance therapy on outcome of dogs with appendicular osteosarcoma following amputation and carboplatin chemotherapy: a multi-institutional study. PLoS One. 2015;10:e0124889-e0124889
159. Sánchez-Céspedes R, Accornero P, Miretti S, Martignani E, Gattino F, Maniscalco L. et al. In vitro and in vivo effects of toceranib phosphate on canine osteosarcoma cell lines and xenograft orthotopic models. Vet Comp Oncol. 2020;18:117-127
160. Schuetze SM, Wathen JK, Lucas DR, Choy E, Samuels BL, Staddon AP. et al. SARC009: Phase 2 study of dasatinib in patients with previously treated, high-grade, advanced sarcoma. Cancer. 2016;122:868-874
161. Crown JP, Diéras V, Staroslawska E, Yardley DA, Bachelot T, Davidson N. et al. Phase III trial of sunitinib in combination with capecitabine versus capecitabine monotherapy for the treatment of patients with pretreated metastatic breast cancer. J Clin Oncol. 2013;31:2870-2878
162. Barrios CH, Liu M-C, Lee SC, Vanlemmens L, Ferrero J-M, Tabei T. et al. Phase III randomized trial of sunitinib versus capecitabine in patients with previously treated HER2-negative advanced breast cancer. Breast Cancer Res Treat. 2010;121:121-131
163. Bergh J, Bondarenko IM, Lichinitser MR, Liljegren A, Greil R, Voytko NL. et al. First-line treatment of advanced breast cancer with sunitinib in combination with docetaxel versus docetaxel alone: results of a prospective, randomized phase III study. J Clin Oncol. 2012;30:921-929
164. Buchbinder EI, Sosman JA, Lawrence DP, McDermott DF, Ramaiya NH, Van den Abbeele AD. et al. Phase 2 study of sunitinib in patients with metastatic mucosal or acral melanoma. Cancer. 2015;121:4007-4015
165. Decoster L, Broek I, Neyns B, Majois F, Baurain JF, Rottey S. et al. Biomarker analysis in a phase II study of sunitinib in patients with advanced melanoma. Anticancer Res. 2015;35:6893-6899
166. Gong HZ, Zheng HY, Li J. The clinical significance of KIT mutations in melanoma: a meta-analysis. Melanoma Res. 2018;28:259-270
167. Meng D, Carvajal RD. KIT as an oncogenic driver in melanoma: an update on clinical development. Am J Clin Dermatol. 2019;20:315-323
168. Murakami A, Mori T, Sakai H, Murakami M, Yanai T, Hoshino Y. et al. Analysis of KIT expression and KIT exon 11 mutations in canine oral malignant melanomas. Vet Comp Oncol. 2011;9:219-224
169. Gomes J, Queiroga FL, Prada J, Pires I. Study of c-kit immunoexpression in canine cutaneous melanocytic tumors. Melanoma Res. 2012;22:195-201
170. Chu P-Y, Pan S-L, Liu C-H, Lee J, Yeh L-S, Liao AT. KIT gene exon 11 mutations in canine malignant melanoma. Vet J. 2013;196:226-230
171. Newman SJ, Jankovsky JM, Rohrbach BW, LeBlanc AK. c-kit expression in canine mucosal melanomas. Vet Pathol. 2011;49:760-765
172. Giuliano A, Dobson J. Prospective clinical trial of masitinib mesylate treatment for advanced stage III and IV canine malignant melanoma. J Small Anim Pract. 2020;61:190-194
173. Londhe P, Gutwillig M, London C. Targeted therapies in veterinary oncology. Vet Clin North Am Small Anim Pract. 2019;49:917-931
174. Cekanova M, Rathore K. Animal models and therapeutic molecular targets of cancer: utility and limitations. Drug Des Devel Ther. 2014;8:1911-1921
175. Paoloni M, Mazcko C, Selting K, Lana S, Barber L, Phillips J. et al. Defining the pharmacodynamic profile and therapeutic index of NHS-IL12 immunocytokine in dogs with malignant melanoma. PLoS One. 2015;10:e0129954
176. Fürdös I, Fazekas J, Singer J, Jensen-Jarolim E. Translating clinical trials from human to veterinary oncology and back. J Transl Med. 2015;13:265-271
177. Hünig T. The storm has cleared: Lessons from the CD28 superagonist TGN1412 trial. Nat Rev Immunol. 2012;12:317-318
178. Liu D, Xiong H, Ellis AE, Northrup NC, Dobbin KK, Shin DM. et al. Canine spontaneous head and neck squamous cell carcinomas represent their human counterparts at the molecular level. PLoS Genet. 2015;11:e1005277
179. Amini P, Nassiri S, Malbon A, Markkanen E. Differential stromal reprogramming in benign and malignant naturally occurring canine mammary tumours identifies disease-modulating stromal components. Sci Rep. 2020;10:5506
Author contact
![]() Corresponding author: E-mail: s.oliveiranl; Tel.: +31-63-410-3460.
Corresponding author: E-mail: s.oliveiranl; Tel.: +31-63-410-3460.