Impact Factor
- Issue 14; 2026
- Issue 13; 2026
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Volume 16; 2026
- Advance Articles
- Past Issues
- Cover Images
- Cover Suggestion
- Index & Coverage
- Special Issues
Introduction
2. Brief characteristics of CNS...
3. Characteristics of most...
4. Supportive roles of brain...
5. Strategies of targeting...
6. Conclusions and perspectives
Acknowledgements
References
International Journal of Biological Sciences
International Journal of Medical Sciences
 Global reach, higher impact
Global reach, higher impact

Theranostics 2020; 10(7):2949-2964. doi:10.7150/thno.40783 This issue Cite
Review
Supportive roles of brain macrophages in CNS metastases and assessment of new approaches targeting their functions
Hua You1, Szymon Baluszek2, Bozena Kaminska1,2 ![]()
1. Affiliated Cancer Hospital & Institute of Guangzhou Medical University, Guangzhou, China
2. Laboratory of Molecular Neurobiology, Nencki Institute of Experimental Biology of the Polish Academy, Warsaw, Poland
Received 2019-9-30; Accepted 2020-1-22; Published 2020-2-10
Abstract

Metastases to the central nervous system (CNS) occur frequently in adults and their frequency increases with the prolonged survival of cancer patients. Patients with CNS metastases have short survival, and modern therapeutics, while effective for extra-cranial cancers, do not reduce metastatic burden. Tumor cells attract and reprogram stromal cells, including tumor-associated macrophages that support cancer growth by promoting tissue remodeling, invasion, immunosuppression and metastasis. Specific roles of brain resident and infiltrating macrophages in creating a pre-metastatic niche for CNS invading cancer cells are less known. There are populations of CNS resident innate immune cells such as: parenchymal microglia and non-parenchymal, CNS border-associated macrophages that colonize CNS in early development and sustain its homeostasis. In this study we summarize available data on potential roles of different brain macrophages in most common brain metastases. We hypothesize that metastatic cancer cells exploit CNS macrophages and their cytoprotective mechanisms to create a pre-metastatic niche and facilitate metastatic growth. We assess current pharmacological strategies to manipulate functions of brain macrophages and hypothesize on their potential use in a therapy of CNS metastases. We conclude that the current data strongly support a notion that microglia, as well as non-parenchymal macrophages and peripheral infiltrating macrophages, are involved in multiple stages of CNS metastases. Understanding their contribution will lead to development of new therapeutic strategies.
Keywords: brain metastases, tumor microenvironment, cancer invasion, intracellular signaling, immune infiltrates, microglia, CNS border associated macrophages, perivascular macrophages, immunosuppression
Introduction
Cancer develops in a heterogeneous tissue microenvironment, and surrounding or infiltrating non-malignant cells play important roles in creating a tumor niche and modulating anti-tumor responses [1]. Tumor-associated macrophages (TAMs) accumulate in many cancers and activate multiple wound healing and tissue repair processes that are frequently associated with local and systemic immunosuppression [2-4]. Several excellent reviews have presented many ways through which TAMs contribute to cancer progression [1-4]. TAMs support tumor invasion by releasing an extracellular matrix (ECM) degrading proteases that contribute to reorganization of the surrounding tissues and facilitate cancer cell invasion. TAMs assist in cancer dissemination via the circulation or the lymphatic system. A majority of circulating cancer cells perish in the circulation, only a fraction of about 0.1% survives and less than 0.01% is capable of forming secondary lesions [5]. Specific features of a small surviving subpopulation of metastasis-initiating cells are a subject of intensive studies. Perivascular macrophages support formation of intravasation sites where cancer cells spread into circulation and promote tumor angiogenesis. Some macrophages may co-migrate with cancer cells to a pre-metastatic niche and promote local remodeling of ECM. The presence of macrophages in the clusters of circulating cancer cells may protect them from the immune system [6].
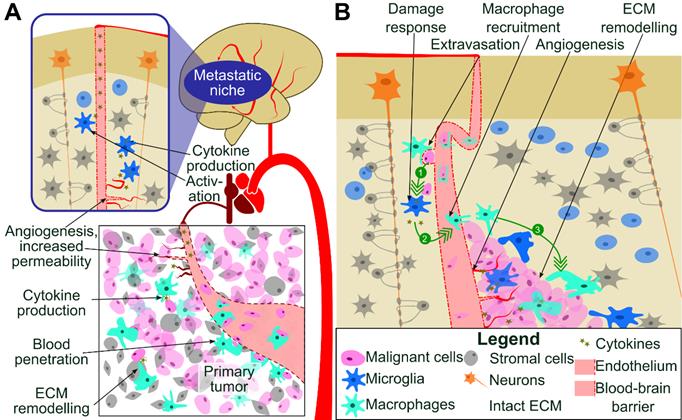
TAMs produce cytokines, chemokines and immunosuppressive molecules that participate in formation of the immunosuppressive microenvironment and contribute to the systemic immunosuppression [4,7]. TAMs-secreted growth factors augment pro-survival pathways and may help tumor cells to resist cytotoxic chemotherapy [8]. Colonization of CNS by cancer cells from a periphery is a complex process involving events such as extravasation from blood vessels, tissue remodeling and death of neurons. All those events result in disturbance of CNS homeostasis and elicit recuperating responses from microglia to protect, repair, and instigate the wound healing, associated with local immunosuppression (Figure 1). All these processes are actively assisted by microglia and infiltrating peripheral macrophages through mechanisms that are poorly characterized [9].
Colonization of CNS by cancer cells from a periphery. (A) Metastatic cancer cells from a periphery colonize CNS via penetration to the blood stream, hematogenous spread, extravasation from blood vessels, seeding a niche followed by tissue remodeling and growth of a secondary cancer. Those processes result in disturbance of CNS homeostasis. (B) Perivascular macrophages and circulating macrophages aid in extravasation of cancer cells (1). Subsequently, metastatic cancer cells secrete cytokines (2) activating CNS resident microglia and infiltrating peripheral macrophages to protect, repair, and instigate tissue repair. These events are associated with local immunosuppression, recruitment of microglia macrophages and tumor growth (3).
2. Brief characteristics of CNS macrophages
CNS is equipped with the resident, innate immune cells called microglia (located in the brain parenchyma) and non-parenchymal, CNS border-associated macrophages (BAMs) consisting of perivascular, meningeal and the choroid plexus macrophages, that are located at the brain-blood vessel interfaces, in the cerebrospinal fluid and in the choroid plexus, respectively. In the past those cells have been collectively called brain macrophages. The genetic lineage tracing approaches and single cell sequencing demonstrated that microglia and BAMs are transcriptionally distinct subpopulations and differ from the bone marrow (BM)-derived macrophages [10-12]. Perivascular macrophages are located in the perivascular space surrounding arteries and veins penetrating deeply into the brain parenchyma, whereas meningeal macrophages are associated with the meninges. Perivascular macrophages express many surface proteins such as CD11b, CD45, CD68, CD115, CD206, CX3CR1, F4/80 and CD163, that are useful markers to identify these cells in flow cytometry and immunocytochemistry. CD163 is a membrane glycoprotein belonging to the scavenger receptor cystein-rich (SRCR) superfamily group B. Perivascular macrophages are positive for the phagocytic cell marker CD68 and the mannose receptor - CD206. In murine CNS, CD206 is strongly expressed on perivascular macrophages, and its expression is weak on infiltrating monocytes and microglia. CD163+ cells are positive for MHC class II antigens and the co-stimulatory molecules such as CD80, CD86, and CD40, suggesting their role in antigen recognition and presentation. CD163+ cells have higher expression of CD45 than microglia [12].
CNS macrophages behave as any other macrophages and function as phagocytic, antigen presenting and cytoprotective cells. Microglia, perivascular and meningeal macrophages originate from yolk sac myeloid progenitors, colonize CNS during early embryogenesis and persist throughout the entire life [13]. Due to different ontogeny, location in CNS and highly specialized functions in a nervous tissue homeostasis and neuronal plasticity, microglia are distinct from peripheral macrophages [14]. Microglia have numerous extensions and actively inspect the brain parenchyma and spinal cord. Microglia detect and remove damaged cells or apoptotic debris by phagocytosis, participate in adequate tuning of neural circuits and contribute to CNS homeostasis [15]. Perivascular macrophages are important for preserving the integrity of the brain-blood barrier (BBB) [12].
Common immunohistochemical markers (Iba1, HLA-DR, Cd68, F4/80) detect all brain macrophages and have not been particularly effective in distinguishing resident microglia from BAMs and monocytes/macrophages invading human tumor tissues under pathological conditions. In flow cytometry, high CD45 expression can differentiate macrophages (Cd11b+CD45high) from microglia (Cd11b+CD45low). Recent studies using single cell sequencing and cell lineage tracing demonstrated that resident microglia are functionally distinct from BM-derived monocytes, which enter CNS under pathological conditions. Distinct transcriptomic profiles have been found in microglia and BM-derived macrophages infiltrating experimental gliomas and brain metastases in mice [16]. The surface molecule CD49D, the α4 subunit of the integrin heterodimer α4β1 and Ly6C (lymphocyte antigen 6 complex, locus C1) have been proposed as good markers for flow cytometry to discriminate microglia and BM-derived macrophages in human brain tumors [16].
Our knowledge regarding heterogeneity and specific functions of brain macrophages within intracranial metastases is limited. Studying microglia-metastatic carcinoma interactions is hampered by: 1) a lack of convenient immunohistochemical markers distinguishing brain macrophages populations, 2) limited availability of patient samples, and 3) scarcity of appropriate animal models to study the microenvironment of CNS metastases. In this study, we describe the role of those cells in CNS invasion and metastatic spread based on available clinical and experimental observations. Several studies reported the accumulation of HLA-DR+ microglia/macrophages in the intracranial metastatic lesions in breast, melanoma, small cell lung, and non-small cell lung cancers [17], but understanding roles the specific subpopulation of brain macrophages play in metastatic seeding of CNS and cancer progression is incomplete. In the Table 1 we summarize conventional markers of main monocytic populations, their role in CNS biology under normal and pathological conditions [10-16], and potential roles in CNS metastases.
Monocytic populations in the central nervous system and CNS metastases
| Population | Markers | Brief description of functions | Role in metastases |
|---|---|---|---|
| Microglia | CD11b, CD68, CD115, CD206, CX3CR1, F4/80, Fcrls++, Iba1+, MerTK, P2ry12++, Siglec-H | In steady state: maintain homeostasis, exhibit a phagocytic activity, have low activity as antigen presenting cells | |
| CD11b, CD45, CD64, CD68, CD115, CD163+/-, CD206, CX3CR1, F4/80, Fcrls+, Iba1++, P2ry12+ | Upon activation: depending on stimuli polarization to an inflammatory of pro-tumorigenic phenotype: morphological changes, phagocytosis and antigen presentation, re-organization of ECM, cytokine and chemokine production | Tumor supporting phenotype degradation of ECM, modulating adaptive immune response, aiding in angiogenesis | |
| CNS border-associated macrophages (BAMs) | CD11b, CD45, CD68, CD115, CD163, CD206, CX3CR1, F4/80 | In steady state: maintain integrity of BBB. Upon pathological activation: modulate BBB integrity and vascular permeability, interact with circulating immune cells, produce reactive oxygen species. | Tumor supporting phenotype: facilitation of recruitment of cancer cells and immune cells via BBB |
| Infiltrating bone marrow-derived macrophages | CD11b, CD45, CD49D, CD64, CD68, CD115, CD163+/-, CD206, CCR2, F4/80, Ly6C | Infiltrate brain parenchyma mainly after breakdown of BBB; immunoregulatory and immunosuppressive functions; cytoprotective activity. | Tumor supporting phenotype: ECM remodeling, immune suppression, enhancement of angiogenesis. |
3. Characteristics of most common CNS metastases
CNS metastases, with incidence of 8.3 to 14.3 per 100,000 people [18], are recognized as the most common intracranial neoplasms and are found in autopsies of 20% cancer patients [19]. The incidence varies between different tumor types; the most frequent are metastases of lung cancer (40-50%), breast cancer (20-30%), melanomas (20-25%), renal carcinomas (10-20%) and gastrointestinal tumors (4-6%). Predominant locations of CNS metastasis are the brain parenchyma and the leptomeninges. The incidence of CNS metastases has been increasing through recent decades due to multiple factors: increasing population age, prolonged survival of patients with primary and secondary advanced cancers, more effective detection of metastases with advancements in imaging techniques [20,21]. New therapeutics targeting oncogenic kinases and immune checkpoint inhibitors increased overall and progression-free survival in patients, including those with brain metastases, but most of them did not effectively reduce metastatic burden and a majority of oncologic patients still die due to dissemination of the disease [22-24].
CNS metastases affect cognitive functions, speech, coordination, behavior, reduce the quality of life and ultimately lead to death. Standard of care for CNS metastases include local surgery, stereotactic radiosurgery or stereotactic fractionated radiotherapy combined with systemic chemotherapy. Despite the advancements in treating patients with metastatic cancer [22,23], a metastatic burden is cause of death in 90% of cancer patients [24]. With modern treatments the median survival of patients with CNS metastases is 6 months [25], but the results vary depending on tumor histology, disease control, patient age and initial responses to a specific therapy [26]. CNS metastases typically have a poor prognosis and patients survive 3-6 months, so consequently patients suffering from CNS metastases are sometimes excluded or are underrepresented in clinical trials of new drugs. Clinical trials in melanoma brain metastases with combined PD-1/CTLA-4 blockade showed ~50% intracranial response rate [27,28]. Moreover, checkpoint inhibitors are considered effective as radiosensitizers in CNS metastases [29].
CNS metastases are relatively uncommon in children with the estimated frequency at 13% in autopsy studies and at 1.5% in clinical studies [30]. CNS is frequently affected in pediatric leukemias and rare in solid tumors, where the occurrence is the highest in germ cell tumors, bone and soft tissue sarcomas [18,30]. For leukemias and lymphomas, CNS invasion is carried by blood and spreads through the arterial and capillary system, or via direct expansion from the cranial bone marrow. The choroid plexus (a tissue with a dense network of capillaries) and connecting veins between the bone marrow and superficial arachnoid is suspected to be a site of cancer cell invasion into CNS [31].
Different cancers have varying propensity to form metastases in CNS. Non-small-cell lung cancer (NSCLC) is a most common in invading brain parenchyma (70% of metastases affect brain) and many patients with stage III or IV cancer (up to 55%) develop brain metastases in the course of the disease [32], with 3-5% patients having leptomeningeal metastases [33]. A histological subgroup of small-cell lung cancer (SCLC) affects 20% of patients and is particularly aggressive. These metastases are treated with carboplatin, etoposide and preventive cranial irradiation. Despite recent advancement with anti-PD1 antibody atezolizumab, prognosis remains poor.
Breast cancer is molecularly classified by the expression of estrogen receptor (ER), progesterone receptor (PR) and human epidermal growth factor receptor-2 (HER-2). The triple-negative breast cancer is the most aggressive form. Its metastases to CNS are discovered in 46% of patients, but this reflects rather the overall metastasis burden than a specific property of this molecular subgroup. HER2+ breast cancers are thought to be particularly prone to cause delayed CNS metastases [34]. In general, CNS metastases of breast cancer prognosticate better than to other sites (13.8 months) and respond to a systemic therapy in up to 80% of cases [35].
Metastases to CNS were found in 10-40% of melanoma patients, with the higher number of metastatic lesions (70- 90%) detected in brains during autopsy. Melanoma cells show some preferences as to location in CNS with the majority located within the frontal lobe (43.5%), less frequently in the cerebellum (8.6%) and rarely found in the hippocampus (<0.1%) [36]. BRAF mutations occur in 40-50% of melanomas and treatments with specific inhibitors (e.g. vemurafenib, dabrafenib) were reported to be effective in a metastatic disease. The presence of BRAF mutation does not affect probability of CNS metastases, but a targeted treatment with vemurafenib decreases such probability [37]. Melanomas are highly immunogenic tumors and checkpoint inhibitors have been very successful [38]. Combining potent BRAF inhibitors with checkpoint inhibitors or stereotactic surgery have extended the therapeutic options for treating the brain metastases from melanoma [38].
Neurologic complications are common in leptomeningeal, epidural and brain parenchyma metastases of non-Hodgkin's lymphomas and are associated with a poor prognosis [39]. Acute lymphoblastic leukemia (ALL) has a marked tendency to metastasize to CNS, it occurs in 5% of patients and ALL relapse in CNS predicts poor outcomes. CNS‐directed therapies such as: cranial irradiation, intrathecal chemotherapy and systemic administration of CNS‐penetrating chemotherapeutics, have reduced the frequency of disease recurrence [40]. Spread of ALL rarely involves the parenchyma and is usually confined to the leptomeninges (lymphomatous meningitis).
Whole-exome sequencing of 86 matched brain metastases, primary tumors, and normal tissue examined if brain metastases harbor distinct genetic alterations from the ones observed in primary tumors. Most of the cases were derived from lung, breast and renal cell carcinomas. While all metastatic and primary sites shared mutational profiles suggesting a common ancestor, in 53% of cases, some alterations were found only in the brain metastases. Detected alterations were associated with the PI3K/AKT/mTOR, CDK, and HER2/EGFR signaling pathways and a sensitivity to pathway specific inhibitors in the brain metastases was proposed. Spatially and temporally separated brain metastasis sites were genetically homogenous, while distal extracranial and lymph node metastases were highly divergent from brain metastases [41]. A recent TCGA (The Cancer Genome Atlas) study interrogating genomics of a tumor-of-origin and its metastasis among thousands samples of 33 tumor types revealed that metastases generally retained the mutational landscape of tumor of origin [42]. It has become clear, however, that one of key features leading to metastasis formation is presence of a (pre)metastatic niche. Primary tumor secretome plays crucial role in this process. Exosomal micro-RNA alters microglia and BBB function which enables cancer invasion [43].
4. Supportive roles of brain macrophages in CNS metastases
4.1. Accumulation of microglia and macrophages in CNS metastases and impact on immune microenvironment
HLA-DR, Iba1 and CD68 are widely used as microglia and macrophage markers in a human tissue. HLA-DR is a heterodimeric cell surface glycoprotein comprised of a 36 kD α (heavy) chain and a 27 kD β (light) chain. It is expressed on microglia, monocytes/macrophages and can be weakly expressed on dendritic cells, B cells, and activated T cells. Iba1 is an ionized calcium binding adaptor molecule 1 and acts as a microglia/macrophage-specific calcium-binding protein with actin-bundling activity that participates in membrane ruffling and phagocytosis. CD68 is a member of the class D scavenger receptors and a glycosylated type I membrane protein that belongs to the lysosome-associated membrane proteins in macrophages. CD68 has been widely used as a pan-macrophage marker, although it can be weakly expressed on endothelial cells. In one of the first studies, the presence of brain macrophages in human CNS metastases was detected by immunohistochemistry (IHC) using an anti-CD68 antibody on paraffin-embedded tissue specimens of a small cohort consisting 17 metastatic tumors, including: lung, breast and clear cell kidney carcinomas. CD68+ macrophages were localized within the tumor tissue, at its periphery and its surroundings. In some cases, strongly stained CD68+ cells were visible in blood vessel walls. Those were likely perivascular macrophages. The study did not report any correlation between the type of tumor and extent of macrophage infiltration [44].
Further studies demonstrated Iba1+ cells with amoeboid, activated morphology found close to the cancer cells in lung cancer CNS metastatic lesions. Double labeling for the inflammatory markers revealed that a majority of those Iba1+ cells did not co-localize with either iNOS (inducible nitric oxide synthase) or TNF-α (tumor necrosis factor-α) staining, which can be interpreted as a lack of inflammatory, antitumor activation, and acquirement of the pro-tumorigenic phenotype [45]. The number of CD68+ cells detected in metastases of lung adenocarcinoma was high and most positive cells displayed amoeboid morphology [46]. Interestingly, the presence of activated brain macrophages correlated with the active immune microenvironment in SCLC brain metastases. Subsets of tumor infiltrating lymphocytes (TILs) such as CD3+, CD8+, CD45RO+, FOXP3+ and PD-1+ cells were analyzed in SCLC brain metastases and four matched primary tumor specimens. Patients with higher numbers of infiltrating CD45RO+ TILS survived longer. The expression of PD-L1 was detected on TILs and on the tumor infiltrating macrophages by immunohistochemistry [47]. Similar proportions of TILs were detected in non-small cell lung carcinoma metastases [48]. Detection of TIL infiltration was similar irrespectively of malignancy degree [48,49], which suggests an active immune microenvironment in those brain metastases.
Staining with the KiM1P antibody (recognizes CD68) showed positive cells in all samples of human breast cancer metastases to CNS and accumulation of amoeboid, positively stained cells in the boundary region between tumor and neighboring tissue. Their number varied from only few up to 50% of all cells [50]. Immunohistochemical staining for a comprehensive panel of 21 inflammation-associated markers, including HLA-DR, in 17 human tissue specimens of brain metastases from breast cancer, NSCLC, SCLC and melanoma showed marked peritumoral accumulation and in some cases intratumoral infiltration of the HLA-DR+ microglia/macrophages. A high proportion of these cells showed a strong immunoreactivity for phagocytosis-associated markers, while a smaller subgroup of cells expressed molecules involved in production of free radicals. Only few B- and T-lymphocytes were observed in and around the brain metastases, and only a fraction of T-cells showed Granzyme B expression. Melanoma brain metastases had significantly less peritumoral brain macrophages than brain metastases of NSCLC did. The authors concluded that microglia/macrophages in brain metastases are activated and up-regulate proteins involved in phagocytosis, but activated cells do not activate the adaptive immunity [17]. Altogether, the results clearly demonstrate accumulation of active (amoeboid) microglia/macrophages in CNS metastases of lung, breast cancers and melanomas. A few observations suggest that those cells did not show signs of the inflammatory, antitumor activation and rather adapt the pro-tumorigenic phenotype.
While most of the metastases were found in the brain parenchyma, metastases to the dura matter, one of the layers that cover the spinal cord and brain (meninges), were found during autopsy in 9-10% of all patients with different cancers [51]. Neoplastic spread to leptomeninges is a result of cancer dissemination to the cerebrospinal fluid, which allows cancer cells to travel to multiple sites within the CNS, extravasate, and grow. Leptomeningeal metastases occur in 1-5% of patients with solid tumors and 5-15% of patients with leukemia [52]. Melanoma, lung and breast cancers may form leptomeningeal metastases and a median survival in such cases is extremely short (2-3 months). Currently, there is no data on accumulation of microglia and macrophages in those tumors, but perivascular and meningeal macrophages due to their location may play a role in CNS metastases, as those cells are involved in immune-surveillance and participate in the recruitment of peripheral immune cells into the CNS in a response to pathological stimuli [53].
Primary central nervous system lymphoma (PCNSL) is a primary tumor but due to its location in CNS and interactions with local immune cells shares certain mechanisms with CNS metastases. The PCNSL is a rare form of lymphoma and accounts for 3%-4% of all primary brain tumors and 4%-6% of extra-nodal lymphomas [54]. The majority of PCNSL is pathologically classified as diffuse large B-cell lymphoma (DLBCL) confined to CNS. While the genomic study of PCNSL samples using whole-exome sequencing and RNA-sequencing showed that PCNSL and DLBCL share some common gene expression and mutation profiles [55], DNA methylation profiles of the PCNSL are different from the systemic DLBCLs or normal lymph nodes, which suggests that PCNSL is a biologically distinct entity from the peripheral DLBCLs [56]. Assessment of the phenotypes of myeloid cells was performed with a small PCNSL cohort (n=43), and numbers of CD68+, CD163+, and CD204+ TAMs were detected but without association with patient's prognosis [57]. An independent study of a PCNSL cohort (n=47) showed the increased numbers of CD68+ cells to be significantly associated with progression-free survival, and a trend was observed for the increased abundance of CD163+ cells and a shorter survival. The IL-10 level in cerebrospinal fluids was correlated with infiltration of CD68 and CD163+ TAMs [58], and the diagnostic and prognostic value of IL10 in the cerebrospinal fluid in PCNSL was confirmed in other studies [59,60]. The recent study, including the largest PCNSL cohort (n=114), showed that the increased number of CD68+ TAMs and indoleamine 2,3-dioxygenase (IDO) positive cells was associated with a favorable prognosis. The increased number of CD204+ cells and a high ratio of CD204+/CD68+ cells, indicative of the tumor-supporting polarization, were associated with a poor prognosis [61]. PD-L1 and IDO1 were overexpressed by macrophage/microglia in PCNSL tissues, and gene expression profiling indicated that IDO1 expression was positively correlated with the expression of macrophage and lymphocyte markers [61]. PD-L1 expression on lymphoma cells correlated positively with the overall survival, whereas PD-L1 expression in the microenvironment showed a negative trend with the overall survival in a PCNSL cohort (n=64) [62]. Osteopontin (encoded by the SPP1 gene) was the most up-regulated gene (~10 fold) in PCNSL compared to non-CNS DLBCL [63]. Osteopontin is a small phosphoprotein and an integrin ligand, which has been implicated in a variety of biological processes such as ECM adhesion and remodeling, cell migration, angiogenesis, proliferation, immunomodulation, chemotaxis of macrophages, dendritic cells, and T cells [64]. Osteopontin may carry numerous interactions with stromal cells. The location of PCNSL cells in the perivascular spaces may indicate interactions of the malignant B cells with components of the blood-brain barrier: endothelial cells with up-regulated MHC class I and II antigens, ICAM-1, and vCAM-1, and perivascular macrophages.
4.2. Roles of microglia and brain macrophages in CNS metastases- lessons from the animal models
Cancer cell metastasis to CNS is modeled in animals by injection of rodent cancer cells (derived from spontaneous, chemically induced and genetically engineered tumors) and human xenografts via a carotid artery or directly to the brain of immunodeficient or immunocompetent rodents [65]. While none of animal models fully reflects tumor progression observed in patients with a metastatic disease, various models provide important mechanistic insights into the metastatic process and allow testing potential therapeutics. Human breast cancer cells of different lines: MDA-MB-435, MDA-MB-231/brain, injected intracardially to immunosuppressed SCID mice and murine 4T1 cells injected to BALB/c mice formed tumors, although cancer cells needed more time to extravasate into the brain parenchyma than into other organs. The cancer cells were cleared from the brain microvessels and extravasated from day 3 to day 7 after injection, with exception of MDA-MB-231/brain cells, which were slower [66]. Cells invading the brain parenchyma induced locally activation of astrocytes and microglia. Astrocytes up-regulated expression of GFAP (glial fibrillary acidic protein), Nestin or both. Microglia (detected with F4/80 staining) infiltrated into the breast cancer mass, accumulated in the surrounding gliosis zone, and formed contacts with cancer cells directly after successful extravasation [66,67]. Microglia associated with cancer cells were heterogeneous and consisted of activated, hypertrophic microglia and reactive microglia with amoeboid cell morphology [66]. When 99LN-BrM cells derived from MMTV:PyMT breast cancer cells were injected to syngeneic, immunocompetent mice with Cx3cr1-based myeloid cells (a lineage tracing model), both infiltrating microglia and BM-derived macrophages were detected in brain metastases [16].
Not only do microglia react to metastasis, it is also crucial for metastatic niche formation. Recently, role of CEMIP (cell migration-inducing and hyaluronan-binding protein) was shown to predict brain metastasis formation. It was selectively produced by cells metastasing to the brain but did not support metastasis growth. It rather upregulated inflammatory response in microglia [68]. Microglia co-cultured with MCF-7 and 410.4 breast cancer cells in vitro and in living brain slice cultures (in which BM‐derived macrophages were absent) promoted cancer cell invasion and colonization of the brain tissue. Blocking microglia function with the bisphosphonate clodronate (an inhibitor of cells of a monocytic lineage) reduced cancer cell invasion. Stimulation of the TLR4 pathway shifted microglia to a pro-inflammatory and anti‐invasive phenotype. In organotypic brain slice cultures, microglia facilitated transport a single invading cell as well as cancer cell cohorts. Gene expression studies of microglia co-cultured with carcinoma cells did not show up-regulation of whole gene-set of the pro-tumorigenic (M2) phenotype but identified TLR and WNT signaling as the most affected pathways in those microglia. Both pathways participate in tissue regeneration and repair [50].
In brain metastases developing after intracarotid injection of different breast cancer cells (4T1, PyMT, or MDA-MB-231), CD11b+F4/80+ cells were detected by flow cytometry as the most abundant infiltrating immune cell population. The infiltration of myeloid-derived suppressor cells (MDSCs; CD11b+Gr1+), granulocytes (CD11b+Ly6G+) and monocytes (CD11b+Ly6C+) into dural metastases was greater than in parenchymal lesions [69]. The T-cells (CD3e+) were rarely detected in either location. Of note, gene expression profiling revealed significant differences in gene expression of cancer cells that have metastasized to the brain parenchyma or the dura, with the high level of mRNA for Lymphotoxin β (LTβ) in parenchymal compared to dural metastatic lesions. The lower levels of inos, MHCII, CD11c, arg1, ifnγ and tnfα in CD11b+ cells from parenchymal versus dural metastasis were detected. The expression of cd206 (a pro-tumorigenic phenotype marker) was significantly increased in parenchymal CD11b+ cells. This pattern of marker gene expression suggests that the parenchymal microglia/macrophages are more twisted towards the pro-tumorigenic phenotype compared to dural cells. It also confirms that a location of a metastatic site matters [69].
Signaling pathways underlying communication of cancer metastatic cells with microglia are similar to the ones operating in peripheral metastases and include Wnt/β‐catenin signaling, CXCR4 and its ligand SDF1 and PI3K pathway. Microglia‐dependent invasion of breast cancer cell co-cultures and in living brain slices was abolished by DKK‐2 (a secreted Wnt antagonist which antagonizes predominantly Wnt/β‐catenin signaling) [50]. The cxcr4 gene coding for C-X-C chemokine receptor 4 (CXCR4) was one of the most up-regulated genes in microglia [50]. CXCR4 and its ligand stroma derived factor 1 (SDF1) are up-regulated in various cancers, and CXCR4 inhibition prevented metastasis formation [70]. Studies of human breast cancer MCF-7 cells and the benign Madin-Darby canine kidney cells (MDCK) injected to brain slices demonstrated that microglia support invasion of breast cancer MCF-7 cells, but not the benign epithelial MDCK cells. The WNT inhibitor DKK2, as well as a CXCR4 inhibitor - AMD3100, reduced invasion of MCF-7 into the whole brain slice with similar efficacy [71]. PI3K signaling was found active in the majority of breast cancer brain metastases. A systematic quantification of the PI3K pathway activity in breast cancer CNS metastases, using a reverse phase protein array, found a high PI3K activation in 62.5% brain metastatic tissues. PI3K signaling was activated in metastasis-promoting microglia/macrophages during CNS colonization. Treatment with buparlisib (BKM120), a pan-PI3K Class I inhibitor, reduced metastasis-promoting activity of microglia/macrophages [72].
Melanoma cells have a different propensity to colonize different organs. When two human melanoma cell lines were injected intracardially to immunodeficient mice: nude (nu/nu), NIH triple immunodeficient (TID: nu/nu, bg/bg, xid/xid) and severe combined immunodeficient (SCID) mice, MM-RU melanoma cells gave rise exclusively to lung metastases, whereas the MM-AN cells gave rise to lung and extra-pulmonary metastases. The metastatic lesions were circumscribed in all organs and had peripherally located macrophages, except for brain metastases, where a more invasive pattern along vasculature was observed [73]. In immunocompetent animals, injected K-1735 melanoma cells formed metastatic lesions only in the brain parenchyma, whereas B16 melanoma cells and mixed B16 x K-1735 melanoma cells formed metastatic lesions only in the leptomeninges and ventricles. The difference in location of a metastatic tumor was likely due to the expression of transforming growth factor-beta 2 (TGF-β2) in melanoma cells: TGF-β2 mRNA was highly expressed by the K-1735 cells, whereas the B16 cells or B16 x K-1735 cell mixes had low expression. Accordingly, manipulation of TGF-β2 expression in melanoma cells reduced metastasis to the brain parenchyma, but did not affect metastasis to the leptomeninges or ventricles [74].
After the intracranial transplantation of spontaneous melanoma brain metastasis cells to immunocompetent mice, activated astrocytes and microglia (stained with isolectin B4, ILB4) were recruited to the tumor-brain interface [75]. Using CX3CR1-GFP transgenic mice allowed the visualization of the dynamic changes of microglia and macrophages during tumor growth of intracranially implanted melanoma cells through intravital imaging. Depletion of microglia and macrophages by treatment with PLX3397, an inhibitor of colony stimulating factor-1 receptor (CSF-1R), reduced the total number and mean size of the brain metastases by 83% and 65%, respectively. Microglia and macrophages from metastatic brains expressed MMP3 and treatment with PD166793, an MMP inhibitor, reduced the total number and mean size of the brain metastases by 50% and 53%, respectively [76]. The results show the supporting role of tumor infiltrating microglia and macrophages.
Preclinical animal models well mimic the clinical course and neuropathology of human PCNSL and show pathological interactions between the malignant B cells, resident cell populations of CNS, and the associated immune infiltrates [77,78]. Those interactions may foster aggressiveness of tumor cells and accelerate the fatal course of disease. A new player in interactions between microglia and cancer cells is Osteopontin (another name SPP1, small secreted phosphoprotein). Osteopontin is an activating factor for microglia and other immune cells. A gene coding for Osteopontin - SPP1 is the most upregulated gene in PCNSL compared to non-CNS DLBCL [63,79]. SPP1 overexpression up-regulates invasiveness of B lymphoma cells in murine brain slices, promotes intracerebral invasion and dissemination of lymphoma cells. It increases the intracerebral lymphoma growth and shortens the survival in athymic mice. Mechanistically, these effects depend on intracellular Osteopontin (iOPN), which is encoded by one of the SPP1 variants. The iOPN acts on transcription factor NFκB and causes transcriptional downregulation of the NF-κB inhibitors, A20/TNFAIP3 and ABIN1/TNIP1, while secretory Osteopontin promotes receptor-mediated activation of NF-κB [63]. It has been shown that glioma-derived Osteopontin is a potent inducer of microglia acting via integrin αvβ3 receptors at the microglial cells [80]. It is likely that Osteopontin, produced by PCNSL cells, will act on microglia stimulating and inducing the pro-tumorigenic phenotype.
4.3. It is all about location - anatomical considerations in CNS metastases
The relative frequency of brain metastases in various anatomical regions of the brain differs: melanoma tends to metastasize to the frontal and temporal lobes, breast carcinoma to the cerebellum and the basal ganglia, large cell carcinoma of the lung to the occipital lobe and squamous cell carcinoma of the lung to the cerebellum [81]. A recent study reported non-uniform distribution of metastatic brain lesions in breast and lung cancer patients. The lesions of NSCL cancer were preferentially located in the parieto-occipital lobes and cerebellum, while breast cancer lesions were in the cerebellum [82]. CNS metastases generally locate in cortical regions, receiving blood supply from the most distal branches of large arteries i.e. anterior, middle and posterior cerebral arteries, and gray-white matter junction, probably due to slow blood flow velocity in those regions. Small intracranial melanoma metastases frequently occur at the interface between the cortex and leptomeninges [83]. Application of high-resolution magnetic resonance imaging to such lesions demonstrated that deeper parenchymal extension of melanoma metastases occurs secondarily and suggests that the leptomeninges are a preferential site for establishment of melanoma metastasis [84]. These findings suggest that different primary tumors could have propensities for different cerebral vascular areas and cerebral edema.
Leptomeningeal spread is a hallmark of hematological malignancies. It occurs in 5-15% of patients with non-Hodgkin's lymphoma and affects up to 10% of patients with acute lymphoblastic leukemia [39,40]. Xenotransplantation of human ALL cells into immunodeficient NSG mice resulted in the infiltration leukemic cells exclusively to meninges and led to development of neurologic symptoms. CXCR4 inhibition (with CXCR4 antagonist AMD‐3100) impaired grafting of ALL cells to bone marrow, leukemia development and CNS infiltration [85]. ALL cells migrate into CNS along vessels that pass directly between bone marrow and the subarachnoid space, bypassing the central circulation. The basement membrane of these vessels is enriched in laminin and its receptor - α6 integrin is expressed on most ALL cells. Interactions between α6 integrin and laminin mediated the migration of ALL cells towards the cerebrospinal fluid. Mice with ALL xenografts treated with a PI3Kδ inhibitor (which decreased α6 integrin expression on ALL cells) or specific α6 integrin-neutralizing antibodies, had significantly reduced transport of ALL cells along vessels [86].
Blood-brain barrier is crucial for maintaining CNS function by protecting neurons from minor disturbances in the systemic homeostasis. BBB is formed by tight junctions of endothelium, pericytes and astrocyte end-feet processes, which should shield CNS from circulating neoplastic cells. However, multiple processes are involved in migration of neoplastic cells through BBB. For instance, expression of Cathepsin S, a member of the cysteine cathepsin protease family, in primary tumor was associated with shorter time to development of brain metastases in immunocompromised mice injected with MDA-MB-231 cells. Of note, Cathepsin S was secreted by both cancer cells and microglia at the metastatic site and likely promoted BBB transmigration through cleavage of junctional proteins. Cathepsin S inhibition in both neoplastic and stromal cells reduced brain metastasis formation [87]. Tumor-secreted micro-RNA were found to promote BBB transmigration. Breast cancer release exosomes containing miR-105 and miR-181c, and transfer their miRNAs to the endothelium which leads to downregulation of tight junctions and disrupts the BBB function [88,89]. Interestingly, the brain microenvironment reciprocates - astrocyte-derived exosomes mediate transfer PTEN-targeting miRNA to metastatic breast cancer cells, leading to downregulation of this tumor suppressor. PTEN loss increases secretion of the chemokine CCL2, which recruits activated microglia [90]. Strategies that could selectively open BBB in the brain metastasis are developed to increase pharmacological efficacy and limit CNS toxicity of the utilized therapy. A novel technique enhancing BBB penetration by pharmaceuticals is combination of focused ultrasounds with intravenously administered microtubules [91]. This approach enhanced both small- and large-molecule drug delivery to the tumor in a murine model of breast cancer metastasis. The enthusiasm for BBB disruption techniques is dampen by the fact that widely used radiotherapy also has BBB-disruptive properties.
5. Strategies of targeting tumor-infiltrating microglia and macrophages in CNS metastases
Recent data on various compounds targeting brain macrophages in different CNS pathologies shed light on their contribution to shaping CNS immune microenvironment. This knowledge can be instrumental in development of potential therapeutics that can be used in a therapy of patients with CNS metastases. Glioblastoma (GBM) is a highly malignant, primary brain tumor which occurs frequently enough to be tested in randomized trials. The supportive role of microglia and macrophages in GBM is well understood and it is fairly well documented how these cells support tumor progression [16]. Moreover, many strategies have been developed recently to target those cells in GBM and some of them have been successful in preclinical studies leading to clinical trials. The lack of drug effectiveness in GBM does not disqualify a specific treatment in CNS metastases and most of those drugs have advantage of crossing effectively BBB.
Experimental studies of CNS metastases in murine models provided a proof of concept for the concept to target tumor infiltrating microglia and macrophages. Application of the bisphosphonate clodronate to brain slice cultures injected with MCF-7 and 410.4 breast cancer cells reduced cancer cell invasion [50]. A critical role of hypoxia in formation of tumor-permissive microenvironment was presented by adding nanoparticles releasing oxygen to culture of M2 macrophages in in vitro model, leading to decreased 4T1 cells mobility and invasiveness. Subsequent reduction of metastatic potential of those cells by the nanoparticles was confirmed in a murine model [92]. Macrophage-colony stimulating factor (CSF-1) signaling through its receptor (CSF-1R) promotes the differentiation of myeloid progenitors into monocytes, macrophages, dendritic cells, and bone-resorbing osteoclasts. CSF-1/CSF-1R signaling facilitates recruitment and survival of TAMs within the tumor microenvironment in many cancers [93]. Pexidartinib (PLX3397) is a novel, orally available, small molecule kinase inhibitor that blocks CSF-1R at an IC50 of 17 nM. The compound has other effects, it inhibits oncogenic, activated FLT3 (FLT3-ITK), interferes with SDF1-induced auto-phosphorylation of c-Kit protein at concentrations below 1 µM and inhibits differentiation of osteoclast precursors in the RANK-L and CSF-1 dependent manner [94]. Treatment with pexidartinib reduced the total number and mean size of the brain metastases of intracranially implanted melanoma cells and improved the efficacy of adoptive cell therapy [95]. Pexidartinib was effective in murine glioma models [96], but did not improve survival when compared with historical controls [97]. A clinical trial of a single agent or combination of CSFR-1 and PD-1 inhibitors in advanced solid tumors is in progress [98].
Several therapeutic strategies have been proposed to target glioma-associated microglia and macrophages. Those compounds target chemokines, metalloproteinases, integrins, tyrosine kinases, interleukin-6, CSF-1 and TGFβ functions, produced or acting both in cancer cells and stromal cells [99]. Unfortunately, a few of those strategies were validated as microglia/macrophages targeting strategies and even fewer were explored in clinical trials. Figure 2 summarizes potential targets for therapeutics targeting microglia.
A scheme illustrating intracellular signaling pathways that could be targeted with existing anticancer drugs in microglia in CNS metastases. Activation of integrins stimulates a focal adhesion kinase (FAK) which forms a complex with Src family kinases, which initiates multiple downstream signaling pathways through phosphorylation of other proteins. Among those pathways are phosphatidylinositol 3-kinase (PI-3K), phosphatidylinositol-4,5-bisphosphate (PIP2), phosphatidylinositol-3,4,5-bisphosphate (PIP3), small G proteins (Rho, Rac, Cdc42), AKT/protein kinase B, ERK (extracellularly regulated kinase) and JNK (Jun N terminal kinase), and mTOR (mammalian target of rapamycin), all regulating different cellular functions. HDACs are histone deacetylases, epigenetic regulators that regulate histones, protein-DNA interaction, chromatin conformation, and transcription.
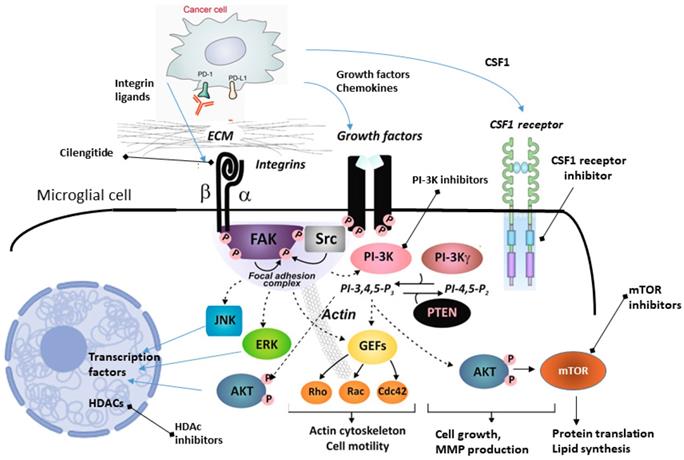
Of interest, cilengitide, a cyclic pentapeptide (cyclo-Arg-Gly-Asp-DPhe-NMe-Val) mimicking the RGD motif binding site, is a selective antagonist αvβ3 and αvβ5 integrin. It has bimodal action on blood vessels and brain tumor cells, inducing a cell death (anoikis, a cell death due to lack of anchorage) in the angiogenic endothelium and glioma cells. Activation of integrin signaling stimulates a focal adhesion kinase (FAK) which forms a complex with Src family kinases, which initiates multiple downstream signaling pathways through phosphorylation of other proteins. Among those pathways are phosphatidylinositol 3-kinase (PI-3K), phosphatidylinositol-4,5-bisphosphate (PIP2), phosphatidylinositol-3,4,5-bisphosphate (PIP3), AKT/protein kinase B, kinase ERK (extracellularly regulated kinase) and JNK (Jun N terminal kinase), and mTOR (mammalian target of rapamycin), all regulating different cellular functions (Figure 2). Interference in ECM or ligands binding to integrins, should affect all downstream signaling pathways.
Cilengitide showed some efficacy in the phase II CORE clinical trial and higher αvβ3 levels in tumor cells were associated with improved progression-free and overall survival [100]. Microglia express high levels of αvβ3 integrins [80,100] and αvβ3 and αvβ5 integrins are expressed on macrophages [100]. However, cilengitide failed to show superiority to temolozomide therapy in gliomas in the randomized phase III CENTRIC trial [101]. Protein S (PROS1), a known ligand of TAM (Tyro-3, Axl, and Mer) receptor tyrosine kinase family [102] is secreted by tumor-associated macrophages/microglia and associates with an activated AXL kinase in mesenchymal glioma stem cells. Interestingly, PROS1 binds to AXL which leads to its activation and downstream targets of AXL, such as NF-κB, are activated, leading to expression of PD-L1 and subsequent cell growth. BGB324, a novel small molecule inhibitor of AXL, alone prolonged the survival of mice bearing human mesenchymal GBM cells, and combinatorial therapy with BGB324 and nivolumab prolonged the survival of mice bearing GBM tumors and increased cell death [103]. AXL expression is an independent prognostic biomarker for survival outcome in brain metastases of NSCLC [104].
There is shortage of data regarding effectiveness of pharmaceuticals specifically affecting microglia/macrophages in brain metastases. Pexidartinib showed promising efficacy in breast cancer [105] and cilengitide was ineffective in melanoma and pancreatic cancer [106,107], but drugs were not explored for intracranial disease activity.
PI-3K is a common component of numerous signaling pathway activated by stimulation of integrins, receptors of growth factors, chemokines and cytokines in both cancer cells and microglia. Activating mutations in genes related to the PI-3K/AKT/mTOR-pathway occur in 43-70% of breast cancer brain metastasis patients. Buparlisib (BKM120) is a specific, oral pan-PI3K class I inhibitor. Treatment with BKM120 reduced metastasis-promoting activity of microglia/macrophages in animal models of breast cancer metastasis [72] and is currently under investigation in patients with breast cancer CNS metastases. The Akt-inhibitor GDC-0068 displayed antitumor activity towards breast cancer metastatic cells with activating mutations in the PI3K pathway in vitro and inhibited the growth of PIK3CA-mutant tumors breast cancer brain metastases in vivo [108]. The inhibitors of mTOR are widely used in renal cell carcinoma, are safe in brain metastases [109] and are explored in combinations specifically for breast cancer [110]. Whether some of their effects are mediated by inhibition of tumor infiltrating microglia is currently unclear. Notably, a recently developed platinum complex with selective activity against cells with active NFκB signaling was shown to act on activated macrophages [111].
There are indications of adding bevacizumab to existing treatment combinations in patients with various solid tumors brain metastases [112-114]. VEGF signaling is involved in TAMs chemoattraction and proliferation [115], therefore some effects could be mediated by inhibiting microglia/macrophages. In a retrospective analysis of three clinical phase III trials, bevacizumab prevented or delayed the formation of brain metastases of non-small cell lung cancer, but not its metastases outside CNS [112]. There was no effect on HER2 negative and HER2 positive breast cancer CNS metastases. In mice, treatment with bevacizumab inhibited formation of brain metastases by NSCLC cells and in improved overall survival [116].
A selective class IIa histone deacetylase (HDAC) inhibitor, TMP195, affected human monocyte responses to macrophage polarizing cytokines CSF-1 and CSF-2 in vitro [117]. In vivo, TMP195 treatment modulated macrophage phenotypes, altered the MMTV-PyMT breast cancer microenvironment, reduced tumor growth and pulmonary metastases. TMP195 induced the recruitment and differentiation of highly phagocytic, activated macrophages. When combined with chemotherapy regimens or T-cell checkpoint blockade, TMP195 treatment significantly reduced tumor burden [118].
There is a rationale to use immune checkpoint inhibitors in CNS metastases. Immune checkpoint inhibitors (anti-CLTA-4 antibodies and anti-PD-1/PD-L1 antibodies) potentiate host antitumor immune response and demonstrated an impressive clinical efficacy in advanced melanoma, metastatic kidney cancer, and metastatic non-small cell lung cancer, all malignancies that frequently cause brain metastases [27-29,38]. Analysis of patients with brain metastases specimens showed that CD3+ TILs are present in 99.1% specimens and 56% cases had dense TIL infiltration. High density of CD3+ TILs was associated with longer median overall survival (15 months versus 6 months, respectively). The highest density of CD3+ TILs, CD8+ TILs, and PD-1-expressing T cells was found in melanoma brain metastases [48]. Checkpoint inhibitors show activity in brain metastases of melanoma [119,120] and are at least safe in lung cancers [121,122]. The combination of anti-PD-1 (nivolumab) and anti-CTLA4 (ipilimumab) therapy has a significant activity for CNS metastatic disease with a relatively low CNS-specific toxicity [119]. In patients with melanoma brain metastases treated with the combination an intracranial response rate was 55% [119]. The study of nivolumab in metastatic renal cell carcinoma (NIVOREN study) demonstrated safety and moderate efficacy of the drug [122]. A phase II trial evaluating pembrolizumab (the anti-PD-1 monoclonal antibody) for patients with leptomeningeal carcinomatosis is ongoing [123]. It is unfortunate that only a fraction of patients with untreated asymptomatic brain metastases (26%) can be enroll in such trials.
The effects on checkpoint inhibitors on accumulation of TAMs are not clear. Combination of PD-1 and CTLA-4 blockade reduced macrophage infiltration in B16 melanomas [103,124]. In a melanoma model with an intracranial and extracranial (subcutaneous) B16 tumor, mimicking the coexistence of metastases inside and outside the brain, intracranial checkpoint inhibitors efficacy was observed only when extracranial tumor was present. The infiltration of microglia (CD11b+F4/80+CD45low) was increased following combined PD-1/CTLA-4 blockade and correlated with intracranial therapeutic efficacy. Simultaneous increase in CSF-1 within tumors was observed, which may explain increased microglia infiltration [125]. Interestingly, repolarization of TAMs using exosome-mimetic nanovesicles derived from pro-inflammatory macrophages (M1NVs) led to release of pro-inflammatory cytokines and induction of antitumor immune responses in vitro and in vivo. Intravenous injection of M1NVs into tumor-bearing mice suppressed tumor growth and in combination with a PD-L1 antibody further reduced the tumor size [126]. The current results of treatments with various inhibitors targeting microglia/macrophages are summarized in the Table 2.
Molecular targets and therapeutics targeting microglia and macrophages
| Molecular target | Affected processes | Pharmaceutical | Trials in glioma | Trials in brain metastases |
|---|---|---|---|---|
| CSF1R | Recruitment and enhancement of cancer invasion [4,103,126] | Pexidartinib BLZ945 | Phase II trial: lacking activity in comparison with historical controls [97] | Limited; the phase I trial showed good activity in breast cancer [105], included in the adaptive phase II clinical trial [127]. |
| αvβ3 and αvβ5 integrins | Microglia-assisted angiogenesis; polarization of microglia [128] | Cilengitide | Phase III trial: lacking activity in comparison with temolozomide [101] | Minimal to none clinical activity in metastatic melanoma and pancreatic cancer [106,107] |
| Immune check point inhibitors | PD-1 or PDL1 CTLA-4 | Nivolumab or/and ipilimumab | Phase III trial: lacking activity in comparison with bevacizumab [129] | Nivolumab + ipilimumab combination is active in melanoma brain metastases [119,120], nivolumab is safe and active in NSCLC and renal cell carcinoma brain metastases [121,122] |
| AXL kinase | AXL kinase regulates PD-1 expression [130] | BGB324 alone or with Nivolumab | Inhibits glioma xenographs in pre-clinical trials [103] | Not tested |
| mTOR | Polarization towards tumor-permissive phenotypes [131] | Everolimus | Phase II trial: lacking activity in comparison with historical controls [132] | Everolimus, lapatynib, and capecytabine combination had some activity in breast cancer brain metastases [110]; safe and effective in renal cell carcinoma brain metastases [109] |
| Temsirolimus | Phase II trial: not superior to temolozomide but phosphorylation of mTORSer2448 can influence response [133] | Safe and effective in renal cell carcinoma brain metastases [109] | ||
| VEGF | TAMs chemotaxis and proliferation [115] | Bevacizumab | Improves PFS but not OS in newly diagnosed glioblastoma [134] | Encouraging results of various combinations in phase II trials with patients with brain metastases from NSCLC, breast cancer and colorectal cancer [112-114] |
| Regorafenib | Improves PFS and OS in previously treated glioblastoma [135] | In phase III trials, regorafenib significantly increased OS, and PFS of patients with metastatic colorectal cancer [136] | ||
| Histone deacetylase | Effects dependent on dose or cell types [137]; TAMs polarization [116] | Vorinostat | Improved OS when compared with historical results, a subgroup of patients with clear benefit [138] | Promising results in preclinical models of triple-negative breast cancer [139] |
| Valproate | Increased survival in observational studies [140] and in comparison with historical cohorts [141] | |||
| Romidepsin | Augmented temozolomide sensitivity in human glioma cells [139]; lacking clinical activity [142] |
PFS, progression free survival; OS, overall survival
Although, the existing data are fragmentary, it is likely that compounds affecting functions of microglia (and other brain macrophages) may affect brain metastases. As brain metastases were up to now investigated from a perspective of a specific metastatic site, it is difficult to assess whether the chemotherapeutics activity was mediated by local or systemic cells and factors.
6. Conclusions and perspectives
Breast, lung, and melanoma cancer cells exhibit a high tendency to invade CNS. The brain parenchyma and the leptomeninges/ventricular system represent two distinct microenvironments in CNS and certain cancer cells preferentially colonize those sites acquiring different features in the process. Survival of cancer cells and invasion of CNS are supported by different brain cell populations [143]. Microglia and other non-parenchymal macrophages of CNS play important roles instigating and supporting metastases. Cancer cells up-regulate specific factors and hijack several mechanisms to polarize microglia and infiltrating peripheral macrophages into tumor supporting cells. The presented, yet fragmentary studies show important and unexplored roles of microglia and non-parenchymal, CNS resident macrophages in CNS metastases. There is a growing number of drugs specifically targeting brain tumor infiltrating microglia/macrophages (Figure 2, Table 2) that could be tested for a future use in therapy and/or prevention of CNS metastases. Those drugs have been pre-selected to pass blood brain barrier and their action on immune components of glioma microenvironment has been demonstrated. Many current approaches target similar oncogenic pathways that are activated in primary tumors, which does not account for metastatic cancer evolution and activation of unique, oncogenic mechanisms. Oncogenic pathways activated in extracerebral and brain metastases frequently differ, as do immune responses in CNS. Moreover, dysregulated pathways are different in each type of cancer. While is not yet clear if responses of brain macrophages are a tumor type specific, it has been demonstrated that activated brain macrophages exploit similar signaling pathways, therefore compounds targeting tumor supportive macrophages could be effective in CNS metastases of many cancer types. Certain pathways (i.e., PI3K signaling pathway) may be uniformly activated in different brain metastases and specific inhibitors could be effective in a large spectrum of CNS lesions. The recent “white paper” which presents the advances in basic science, translational, and clinical research in melanoma CNS metastasis and therapeutic management of patients, provides excellent recommendations for making significant clinical impact [144].
Leptomeningeal disease remains a unique challenge due to not fully understood pathobiology and a lack of cellular or animal models. Inclusion of cohorts of these patients to melanoma brain metastasis trials, or separate trials for these patients, will be important to move forward. Evaluating intrathecal therapies in these patients is justified. There is also a need to develop preclinical models of leptomeningeal disease to accelerate rational therapeutic development.
The presence of the blood-brain barrier is a limiting factor for the access of chemotherapeutics to metastatic lesions and several strategies to overcome this barrier have been developed. A number of small molecules crossing into the brain parenchyma, novel formulations of existing chemotherapeutics, and disruptive BBB techniques, including transcranial focused ultrasound coupled with intravenously delivered microbubbles, hyperosmotic agents (i.e. mannitol), radiation-induced permeability, have been reported [145]. Basic research and preclinical studies concentrated specifically on brain metastases can lead to further improvements in systemic therapies for these patients.
Acknowledgements
Funding
The research in the laboratory of Dr. Hua You is funded by National Natural Science Foundation of China (81911530169, 81903088, 81670180, and 81711540047).
Author Contributions
HU, SB, BK conceived the hypothesis, did the literature search, and wrote the manuscript; SB and BK prepared the figures.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Hanahan D, Coussens LM. Accessories to the Crime: Functions of Cells Recruited to the Tumor Microenvironment. Cancer Cell. 2012;21:309-22
2. Muliaditan T, Caron J, Okesola M, Opzoomer JW, Kosti P, Georgouli M. et al. Macrophages are exploited from an innate wound healing response to facilitate cancer metastasis. Nat Commun. 2018;9:2951
3. Grivennikov SI, Greten FR, Karin M. Immunity, Inflammation, and Cancer. Cell. 2010;140:883-99
4. Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature. 2008;454:436-44
5. Krebs MG, Hou JM, Ward TH, Blackhall FH, Dive C. Circulating tumour cells: Their utility in cancer management and predicting outcomes. Ther Adv Med Oncol. 2010;2:351-65
6. Hamilton G, Rath B. Circulating tumor cell interactions with macrophages: implications for biology and treatment. Transl Lung Cancer Res. 2017;6:418-30
7. Seike T, Fujita K, Yamakawa Y, Kido MA, Takiguchi S, Teramoto N. et al. Interaction between lung cancer cells and astrocytes via specific inflammatory cytokines in the microenvironment of brain metastasis. Clin Exp Metastasis. 2011;28:13-25
8. Sanchez LR, Borriello L, Entenberg D, Condeelis JS, Oktay MH, Karagiannis GS. The emerging roles of macrophages in cancer metastasis and response to chemotherapy. J Leukoc Biol. 2019;106:259-74
9. Berghoff AS, Preusser M. The inflammatory microenvironment in brain metastases: Potential treatment target? Chinese Clin Oncol. 2015;4:21
10. Prinz M, Priller J, Sisodia SS, Ransohoff RM. Heterogeneity of CNS myeloid cells and their roles in neurodegeneration. Nat Neurosci. 2011;14:1227-35
11. Goldmann T, Wieghofer P, Jordão MJC, Prutek F, Hagemeyer N, Frenzel K. et al. Origin, fate and dynamics of macrophages at central nervous system interfaces. Nat Immunol. 2016;17:797-805
12. Faraco G, Park L, Anrather J, Iadecola C. Brain perivascular macrophages: characterization and functional roles in health and disease. J Mol Med. 2017;95:1143-52
13. Ginhoux F, Lim S, Hoeffel G, Low D, Huber T. Origin and differentiation of microglia. Front Cell Neurosci. 2013:7
14. Butovsky O, Jedrychowski MP, Moore CS, Cialic R, Lanser AJ, Gabriely G. et al. Identification of a unique TGF-β-dependent molecular and functional signature in microglia. Nat Neurosci. 2014;17:131-43
15. Prinz M, Priller J. Microglia and brain macrophages in the molecular age: From origin to neuropsychiatric disease. Nat Rev Neurosci. 2014;15:300-12
16. Bowman RL, Klemm F, Akkari L, Pyonteck SM, Sevenich L, Quail DF. et al. Macrophage Ontogeny Underlies Differences in Tumor-Specific Education in Brain Malignancies. Cell Rep. 2016;17:2445-59
17. Berghoff AS, Lassmann H, Preusser M, Höftberger R. Characterization of the inflammatory response to solid cancer metastases in the human brain. Clin Exp Metastasis. 2013;30:69-81
18. Nayak L, Lee EQ, Wen PY. Epidemiology of brain metastases. Curr Oncol Rep. 2012;14:48-54
19. Budczies J, von Winterfeld M, Klauschen F, Bockmayr M, Lennerz JK, Denkert C. et al. The landscape of metastatic progression patterns across major human cancers. Oncotarget. 2015;6:570-83
20. Tabouret E, Chinot O, Metellus P, Tallet A, Viens P, Gonçalves A. Recent trends in epidemiology of brain metastases: An overview. Anticancer Res. 2012;32:4655-62
21. Giordano SH, Buzdar AU, Smith TL, Kau SW, Yang Y, Hortobagyi GN. Is Breast Cancer Survival Improving? Trends in Survival for Patients with Recurrent Breast Cancer Diagnosed from 1974 through 2000. Cancer. 2004;100:44-52
22. Von Hoff DD, Ervin T, Arena FP, Chiorean EG, Infante J, Moore M. et al. Increased Survival in Pancreatic Cancer with nab-Paclitaxel plus Gemcitabine. N Engl J Med. 2013;369:1691-703
23. Abraham J. Abiraterone increases survival in metastatic prostate cancer. Community Oncol. 2012;9:240-2
24. Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science (80- ). 2011;331:1559-64
25. Stelzer K. Epidemiology and prognosis of brain metastases. Surg Neurol Int. 2013;4:192
26. Hatiboglu MA, Chang EL, Suki D, Sawaya R, Wildrick DM, Weinberg JS. Outcomes and prognostic factors for patients with brainstem metastases undergoing stereotactic radiosurgery. Neurosurgery. 2011;69:796-806
27. Iorgulescu B, Harary M, Zogg C, Ligon KL, Reardon DA, Hodi FS. et al. Risk-adjusted survival for melanoma brain metastases in the era of checkpoint blockade immunotherapies: Results from a national cohort. J Clin Oncol. 2018;36:2011-2011
28. Sloot S, Chen YA, Zhao X, Weber JL, Benedict JJ, Mulé JJ. et al. Improved survival of patients with melanoma brain metastases in the era of targeted BRAF and immune checkpoint therapies. Cancer. 2018;124:297-305
29. Ahmed KA, Stallworth DG, Kim Y, Johnstone PAS, Harrison LB, Caudell JJ. et al. Clinical outcomes of melanoma brain metastases treated with stereotactic radiation and anti-PD-1 therapy. Ann Oncol. 2016;27:434-41
30. Graus F, Walker RW, Allen JC. Brain metastases in children. J Pediatr. 1983;103:558-61
31. Wilhelm I, Fazakas C, Molnár K, Végh AG, Haskó J, Krizbai IA. Foe or friend? Janus-faces of the neurovascular unit in the formation of brain metastases. J Cereb Blood Flow Metab. 2018;38:563-87
32. Chen AM, Jahan TM, Jablons DM, Garcia J, Larson DA. Risk of cerebral metastases and neurological death after pathological complete response to neoadjuvant therapy for locally advanced nonsmall-cell lung cancer: Clinical implications for the subsequent management of the brain. Cancer. 2007;109:1668-75
33. Levy A, Faivre-Finn C, Hasan B, De Maio E, Berghoff AS, Girard N. et al. Diversity of brain metastases screening and management in non-small cell lung cancer in Europe: Results of the European Organisation for Research and Treatment of Cancer Lung Cancer Group survey. Eur J Cancer. 2018;93:37-46
34. Lin NU, Claus E, Sohl J, Razzak AR, Arnaout A, Winer EP. Sites of distant recurrence and clinical outcomes in patients with metastatic triple-negative breast cancer: High incidence of central nervous system metastases. Cancer. 2008;113:2638-45
35. Sperduto PW, Kased N, Roberge D, Xu Z, Shanley R, Luo X. et al. Summary report on the graded prognostic assessment: An accurate and facile diagnosis-specific tool to estimate survival for patients with brain metastases. J Clin Oncol. 2012;30:419-25
36. Gummadi T, Zhang BY, Valpione S, Kim C, Kottschade LA, Mittapalli RK. et al. Impact of BRAF mutation and BRAF inhibition on melanoma brain metastases. Melanoma Res. 2015;25:75-9
37. Davies MA, Saiag P, Robert C, Grob JJ, Flaherty KT, Arance A. et al. Dabrafenib plus trametinib in patients with BRAFV600-mutant melanoma brain metastases (COMBI-MB): a multicentre, multicohort, open-label, phase 2 trial. Lancet Oncol. 2017;18:863-73
38. Larkin J, Chiarion-Sileni V, Gonzalez R, Grob JJ, Cowey CL, Lao CD. et al. Combined nivolumab and ipilimumab or monotherapy in untreated Melanoma. N Engl J Med. 2015;373:23-34
39. Grier J, Batchelor T. Metastatic neurologic complications of non-Hodgkin's lymphoma. Curr Oncol Rep. 2005;7:55-60
40. Clarke M, Gaynon P, Hann I, Harrison G, Masera G, Peto R. et al. CNS-directed therapy for childhood acute lymphoblastic leukemia: Childhood ALL Collaborative Group overview of 43 randomized trials. J Clin Oncol. 2003;21:1798-809
41. Brastianos PK, Carter SL, Santagata S, Cahill DP, Taylor-Weiner A, Jones RT. et al. Genomic Characterization of Brain Metastases Reveals Branched Evolution and Potential Therapeutic Targets. Cancer Discov. 2015;5:1164-77
42. Zehir A, Benayed R, Shah RH, Syed A, Middha S, Kim HR. et al. Mutational landscape of metastatic cancer revealed from prospective clinical sequencing of 10,000 patients. Nat Med. 2017;23:703-13
43. Carvalho R, Paredes J, Ribeiro AS. Impact of breast cancer cells´ secretome on the brain metastatic niche remodeling. Semin Cancer Biol. 2019
44. Kałuza J. The monoclonal antibody (MAB) CD 68 allows the immunocytochemical identification of macrophages in primary and metastatic brain tumors in paraffin embedded tissues. Folia Histochem Cytobiol. 1992;30:125-7
45. He BP, Wang JJ, Zhang X, Wu Y, Wang M, Bay BH. et al. Differential reactions of microglia to brain metastasis of lung cancer. Mol Med. 2006;12:161-70
46. Strik HM, Stoll M, Meyermann R. Immune Cell Infiltration of Intrinsic and Metastatic Intracranial Tumours. Anticancer Res. 2004;24:37-42
47. Berghoff AS, Ricken G, Wilhelm D, Rajky O, Widhalm G, Dieckmann K. et al. Tumor infiltrating lymphocytes and PD-L1 expression in brain metastases of small cell lung cancer (SCLC). J Neurooncol. 2016;130:19-29
48. Berghoff AS, Fuchs E, Ricken G, Mlecnik B, Bindea G, Spanberger T. et al. Density of tumor-infiltrating lymphocytes correlates with extent of brain edema and overall survival time in patients with brain metastases. Oncoimmunology. 2016;5:e1057388
49. El Rassy E, Botticella A, Kattan J, Le Péchoux C, Besse B, Hendriks L. Non-small cell lung cancer brain metastases and the immune system: From brain metastases development to treatment. Cancer Treat Rev. 2018;68:69-79
50. Pukrop T, Dehghani F, Chuang HN, Lohaus R, Bayanga K, Heermann S. et al. Microglia promote colonization of brain tissue by breast cancer cells in a Wnt-dependent way. Glia. 2010;58:1477-89
51. Nayak L, Abrey LE, Iwamoto FM. Intracranial dural metastases. Cancer. 2009;115:1947-53
52. Taillibert S, Chamberlain MC. Leptomeningeal metastasis. Handb Clin Neurol. 2018;149:3-23
53. Bechmann I, Priller J, Kovac A, Böntert M, Wehner T, Klett FF. et al. Immune surveillance of mouse brain perivascular spaces by blood-borne macrophages. Eur J Neurosci. 2001;14:1651-8
54. Kasenda B, Ferreri AJM, Marturano E, Forst D, Bromberg J, Ghesquieres H. et al. First-line treatment and outcome of elderly patients with primary central nervous system lymphoma (PCNSL)-a systematic review and individual patient data meta-analysis. Ann Oncol. 2015;26:1305-13
55. Fukumura K, Kawazu M, Kojima S, Ueno T, Sai E, Soda M. et al. Genomic characterization of primary central nervous system lymphoma. Acta Neuropathol. 2016;131:865-75
56. Nakamura T, Yamashita S, Fukumura K, Nakabayashi J, Tanaka K, Tamura K. et al. Genome-wide DNA methylation profiling identifies primary central nervous system lymphoma as a distinct entity different from systemic diffuse large B-cell lymphoma. Acta Neuropathol. 2017;133:321-4
57. Komohara Y, Horlad H, Ohnishi K, Ohta K, Makino K, Hondo H. et al. M2 macrophage/microglial cells induce activation of Stat3 in primary central nervous system lymphoma. J Clin Exp Hematop. 2011;51:93-9
58. Sasayama T, Tanaka K, Mizowaki T, Nagashima H, Nakamizo S, Tanaka H. et al. Tumor-Associated Macrophages Associate with Cerebrospinal Fluid Interleukin-10 and Survival in Primary Central Nervous System Lymphoma (PCNSL). Brain Pathol. 2016;26:479-87
59. Song Y, Zhang W, Zhang L, Wu W, Zhang Y, Han X. et al. Cerebrospinal fluid IL-10 and IL-10/IL-6 as accurate diagnostic biomarkers for primary central nervous system large B-cell lymphoma. Sci Rep. 2016;6:38671
60. Sasayama T, Nakamizo S, Nishihara M, Kawamura A, Tanaka H, Mizukawa K. et al. Cerebrospinal fluid interleukin-10 is a potentially useful biomarker in immunocompetent primary central nervous system lymphoma (PCNSL). Neuro Oncol. 2012;14:368-80
61. Nam SJ, Kim S, Kwon D, Kim H, Kim S, Lee E. et al. Prognostic implications of tumor-infiltrating macrophages, M2 macrophages, regulatory T-cells, and indoleamine 2,3-dioxygenase-positive cells in primary diffuse large B-cell lymphoma of the central nervous system. Oncoimmunology. 2018;7:e1442164
62. Hayano A, Komohara Y, Takashima Y, Takeya H, Homma J, Fukai J. et al. Programmed cell death ligand 1 expression in primary central nervous system lymphomas: A clinicopathological study. Anticancer Res. 2017;37:5655-66
63. Yushi Q, Li Z, Von Roemeling CA, Doeppler H, Marlow LA, Kim BYS. et al. Osteopontin is a multi-faceted pro-tumorigenic driver for central nervous system lymphoma. Oncotarget. 2016;7:32156-71
64. Anborgh PH, Mutrie JC, Tuck AB, Chambers AF. Role of the metastasis-promoting protein osteopontin in the tumour microenvironment. J Cell Mol Med. 2010;14:2037-44
65. Daphu I, Sundstrøm T, Horn S, Huszthy PC, Niclou SP, Sakariassen PØ. et al. In vivo animal models for studying brain metastasis: value and limitations. Clin Exp Metastasis. 2013;30:695-710
66. Lorger M, Felding-Habermann B. Capturing changes in the brain microenvironment during initial steps of breast cancer brain metastasis. Am J Pathol. 2010;176:2958-71
67. Fitzgerald DP, Palmieri D, Hua E, Hargrave E, Herring JM, Qian Y. et al. Reactive glia are recruited by highly proliferative brain metastases of breast cancer and promote tumor cell colonization. Clin Exp Metastasis. 2008;25:799-810
68. Rodrigues G, Hoshino A, Kenific CM, Matei IR, Steiner L, Freitas D. et al. Tumour exosomal CEMIP protein promotes cancer cell colonization in brain metastasis. Nat Cell Biol. 2019;21:1403-12
69. Rippaus N, Taggart D, Williams J, Andreou T, Wurdak H, Wronski K. et al. Metastatic site-specific polarization of macrophages in intracranial breast cancer metastases. Oncotarget. 2016;7:41473-87
70. Zlotnik A, Burkhardt AM, Homey B. Homeostatic chemokine receptors and organ-specific metastasis. Nat Rev Immunol. 2011;11:597-606
71. Chuang HN, van Rossum D, Sieger D, Siam L, Klemm F, Bleckmann A. et al. Carcinoma cells misuse the host tissue damage response to invade the brain. Glia. 2013;61:1331-46
72. Blazquez R, Wlochowitz D, Wolff A, Seitz S, Wachter A, Perera-Bel J. et al. PI3K: A master regulator of brain metastasis-promoting macrophages/microglia. Glia. 2018;66:2438-55
73. Byers HR, Etoh T, Lee KW, Mihm MC, Gattoni-Celli S. Organ-specific metastases in immunodeficient mice injected with human melanoma cells: A quantitative pathological analysis. Melanoma Res. 1993;3:247-53
74. Zhang C, Zhang F, Tsan R, Fidler IJ. Transforming growth factor-β2 is a molecular determinant for site-specific melanoma metastasis in the brain. Cancer Res. 2009;69:828-35
75. Schwartz H, Blacher E, Amer M, Livneh N, Abramovitz L, Klein A. et al. Incipient melanoma brain metastases instigate astrogliosis and neuroinflammation. Cancer Res. 2016;76:4359-71
76. Qiao S, Qian Y, Xu G, Luo Q, Zhang Z. Long-term characterization of activated microglia/macrophages facilitating the development of experimental brain metastasis through intravital microscopic imaging. J Neuroinflammation. 2019;16:4
77. Deckert M, Montesinos-Rongen M, Brunn A, Siebert R. Systems biology of primary CNS lymphoma: From genetic aberrations to modeling in mice. Acta Neuropathol. 2014;127:175-88
78. Ben Abdelwahed Bagga R, Donnou S, Cosette J, Sautès-Fridman C, Aouni M, Fisson S. Mouse models of primary central nervous system lymphomas: tools for basing funding and therapeutic strategies. J Neurooncol. 2015;121:9-18
79. Rubenstein JL, Shen A, Batchelor TT, Kadoch C, Treseler P, Shuman MA. Differential gene expression in central nervous system lymphoma. Blood. 2009;113:266-7
80. Ellert-Miklaszewska A, Wisniewski P, Kijewska M, Gajdanowicz P, Pszczolkowska D, Przanowski P. et al. Tumour-processed osteopontin and lactadherin drive the protumorigenic reprogramming of microglia and glioma progression. Oncogene. 2016;35:6366-77
81. Graf A-H, Buchberger W, Langmayr H, Schmid KW. Site preference of metastatic tumours of the brain. Virchows Arch A Pathol Anat Histopathol. 1988;412:493-8
82. Quattrocchi CC, Errante Y, Gaudino C, Mallio CA, Giona A, Santini D. et al. Spatial brain distribution of intra-axial metastatic lesions in breast and lung cancer patients. J Neurooncol. 2012;110:79-87
83. Mampre D, Ehresman J, Alvarado-Estrada K, Wijesekera O, Sarabia-Estrada R, Quinones-Hinojosa A. et al. Propensity for different vascular distributions and cerebral edema of intraparenchymal brain metastases from different primary cancers. J Neurooncol. 2019;143:115-22
84. Lasocki A, Khoo C, Lau PKH, Kok DL, Mcarthur GA. High-resolution MRI demonstrates that more than 90% of small intracranial melanoma metastases develop in close relationship to the leptomeninges. Neuro Oncol. 2019
85. Jost TR, Borga C, Radaelli E, Romagnani A, Perruzza L, Omodho L. et al. Role of CXCR4-mediated bone marrow colonization in CNS infiltration by T cell acute lymphoblastic leukemia. J Leukoc Biol. 2016;99:1077-87
86. Yao H, Price TT, Cantelli G, Ngo B, Warner MJ, Olivere L. et al. Leukaemia hijacks a neural mechanism to invade the central nervous system. Nature. 2018;560:55-60
87. Sevenich L, Bowman RL, Mason SD, Quail DF, Rapaport F, Elie BT. et al. Analysis of tumour- and stroma-supplied proteolytic networks reveals a brain-metastasis-promoting role for cathepsin S. Nat Cell Biol. 2014;16:876-88
88. Zhou W, Fong MY, Min Y, Somlo G, Liu L, Palomares MR. et al. Cancer-Secreted miR-105 Destroys Vascular Endothelial Barriers to Promote Metastasis. Cancer Cell. 2014;25:501-15
89. Tominaga N, Kosaka N, Ono M, Katsuda T, Yoshioka Y, Tamura K. et al. Brain metastatic cancer cells release microRNA-181c-containing extracellular vesicles capable of destructing blood-brain barrier. Nat Commun. 2015;6:6716
90. Zhang L, Zhang S, Yao J, Lowery FJ, Zhang Q, Huang W-C. et al. Microenvironment-induced PTEN loss by exosomal microRNA primes brain metastasis outgrowth. Nature. 2015;527:100-4
91. Arvanitis CD, Askoxylakis V, Guo Y, Datta M, Kloepper J, Ferraro GB. et al. Mechanisms of enhanced drug delivery in brain metastases with focused ultrasound-induced blood-tumor barrier disruption. Proc Natl Acad Sci. 2018;115:E8717-26
92. Zhang H, Zhang X, Ren Y, Cao F, Hou L, Zhang Z. An in situ microenvironmental nano-regulator to inhibit the proliferation and metastasis of 4T1 tumor. Theranostics. 2019;9:3580-94
93. Stanley ER, Chitu V. CSF-1 Receptor Signaling in Myeloid Cells. Cold Spring Harb Perspect Biol. 2014;6:a021857-a021857
94. Cannarile MA, Weisser M, Jacob W, Jegg AM, Ries CH, Rüttinger D. Colony-stimulating factor 1 receptor (CSF1R) inhibitors in cancer therapy. J Immunother Cancer. 2017;5:53
95. Mok S, Koya RC, Tsui C, Xu J, Robert L, Wu L. et al. Inhibition of CSF-1 Receptor Improves the Antitumor Efficacy of Adoptive Cell Transfer Immunotherapy. Cancer Res. 2014;74:153-61
96. Pyonteck SM, Akkari L, Schuhmacher AJ, Bowman RL, Sevenich L, Quail DF. et al. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat Med. 2013;19:1264-72
97. Butowski N, Colman H, De Groot JF, Omuro AM, Nayak L, Wen PY. et al. Orally administered colony stimulating factor 1 receptor inhibitor PLX3397 in recurrent glioblastoma: an Ivy Foundation Early Phase Clinical Trials Consortium phase II study. Neuro Oncol. 2016;18:557-64
98. Novartis Pharmaceuticals. Phase I/II Study of BLZ945 Single Agent or BLZ945 in Combination With PDR001 in Advanced Solid Tumors. <. https://clinicaltrials.gov/ct2/show/NCT02829723>
99. Poon CC, Sarkar S, Yong VW, Kelly JJP. Glioblastoma-associated microglia and macrophages: targets for therapies to improve prognosis. Brain. 2017;140:1548-60
100. Weller M, Nabors LB, Gorlia T, Leske H, Rushing E, Bady P. et al. Cilengitide in newly diagnosed glioblastoma: Biomarker expression and outcome. Oncotarget. 2016;7:15018-32
101. Stupp R, Hegi ME, Gorlia T, Erridge SC, Perry J, Hong Y-K. et al. Cilengitide combined with standard treatment for patients with newly diagnosed glioblastoma with methylated MGMT promoter (CENTRIC EORTC 26071-22072 study): a multicentre, randomised, open-label, phase 3 trial. Lancet Oncol. 2014;15:1100-8
102. Linger RMA, Keating AK, Earp HS, Graham DK. TAM Receptor Tyrosine Kinases: Biologic Functions, Signaling, and Potential Therapeutic Targeting in Human Cancer. In. 2008:35-83
103. Sadahiro H, Kang KD, Gibson JT, Minata M, Yu H, Shi J. et al. Activation of the receptor tyrosine kinase AXL regulates the immune microenvironment in glioblastoma. Cancer Res. 2018;78:3002-13
104. Wu X, Ma W, Zhou Q, Yan H, Lim Z-F, Huang M. et al. AXL-GAS6 expression can predict for adverse prognosis in non-small cell lung cancer with brain metastases. J Cancer Res Clin Oncol. 2017;143:1947-57
105. Wesolowski R, Sharma N, Reebel L, Rodal MB, Peck A, West BL. et al. Phase Ib study of the combination of pexidartinib (PLX3397), a CSF-1R inhibitor, and paclitaxel in patients with advanced solid tumors. Ther Adv Med Oncol. 2019;11:175883591985423
106. Kim KB, Prieto V, Joseph RW, Diwan AH, Gallick GE, Papadopoulos NE. et al. A randomized phase II study of cilengitide (EMD 121974) in patients with metastatic melanoma. Melanoma Res. 2012;22:294-301
107. Friess H, Langrehr JM, Oettle H, Raedle J, Niedergethmann M, Dittrich C. et al. A randomized multi-center phase II trial of the angiogenesis inhibitor Cilengitide (EMD 121974) and gemcitabine compared with gemcitabine alone in advanced unresectable pancreatic cancer. BMC Cancer. 2006;6:285
108. Ippen FM, Grosch JK, Subramanian M, Kuter BMF, Liederer BM, Plise EG. et al. Targeting the PI3K/Akt/mTOR pathway with the pan-Akt inhibitor GDC-0068 in PIK3CA-mutant breast cancer brain metastases. Neuro Oncol. 2019;21:1401-11
109. Bassanelli M, Viterbo A, Roberto M, Giacinti S, Staddon A, Aschelter AM. et al. Multimodality treatment of brain metastases from renal cell carcinoma in the era of targeted therapy. Ther Adv Med Oncol. 2016;8:450-9
110. Van Swearingen AED, Siegel MB, Deal AM, Sambade MJ, Hoyle A, Hayes DN. et al. LCCC 1025: a phase II study of everolimus, trastuzumab, and vinorelbine to treat progressive HER2-positive breast cancer brain metastases. Breast Cancer Res Treat. 2018;171:637-48
111. Zhu Y, Zhang M, Luo L, Gill MR, De Pace C, Battaglia G. et al. NF-κB hijacking theranostic Pt(ll) complex in cancer therapy. Theranostics. 2019;9:2158-66
112. Besse B, Le Moulec S, Mazieres J, Senellart H, Barlesi F, Chouaid C. et al. Bevacizumab in Patients with Nonsquamous Non-Small Cell Lung Cancer and Asymptomatic, Untreated Brain Metastases (BRAIN): A Nonrandomized, Phase II Study. Clin Cancer Res. 2015;21:1896-903
113. Lu Y-S, Chen TW-W, Lin C-H, Yeh D-C, Tseng L-M, Wu P-F. et al. Bevacizumab Preconditioning Followed by Etoposide and Cisplatin Is Highly Effective in Treating Brain Metastases of Breast Cancer Progressing from Whole-Brain Radiotherapy. Clin Cancer Res. 2015;21:1851-8
114. Finkelmeier F, You S-J, Waidmann O, Wolff R, Zeuzem S, Bähr O. et al. Bevacizumab in Combination with Chemotherapy for Colorectal Brain Metastasis. J Gastrointest Cancer. 2016;47:82-8
115. Forstreuter F, Lucius R, Mentlein R. Vascular endothelial growth factor induces chemotaxis and proliferation of microglial cells. J Neuroimmunol. 2002;132:93-8
116. Ilhan-Mutlu A, Osswald M, Liao Y, Gömmel M, Reck M, Miles D. et al. Bevacizumab prevents brain metastases formation in lung adenocarcinoma. Mol Cancer Ther. 2016;15:702-10
117. Lobera M, Madauss KP, Pohlhaus DT, Wright QG, Trocha M, Schmidt DR. et al. Selective class IIa histone deacetylase inhibition via a nonchelating zinc-binding group. Nat Chem Biol. 2013;9:319-25
118. Guerriero JL, Sotayo A, Ponichtera HE, Castrillon JA, Pourzia AL, Schad S. et al. Class IIa HDAC inhibition reduces breast tumours and metastases through anti-tumour macrophages. Nature. 2017;543:428-32
119. Long G V, Atkinson V, Lo S, Sandhu S, Guminski AD, Brown MP. et al. Combination nivolumab and ipilimumab or nivolumab alone in melanoma brain metastases: a multicentre randomised phase 2 study. Lancet Oncol. 2018;19:672-81
120. Tawbi HA, Forsyth PA, Algazi A, Hamid O, Hodi FS, Moschos SJ. et al. Combined Nivolumab and Ipilimumab in Melanoma Metastatic to the Brain. N Engl J Med. 2018;379:722-30
121. Bidoli P, Chiari R, Catino A, Grossi F, Noberasco C, Gelsomino F. et al. Efficacy and safety data from patients with advanced squamous NSCLC and brain metastases participating in the nivolumab Expanded Access Programme (EAP) in Italy. Ann Oncol. 2016:27
122. Escudier BJ, Chabaud S, Borchiellini D, Gravis G, Chevreau C, Brachet PE. et al. Efficacy and safety of nivolumab in patients with metastatic renal cell carcinoma (mRCC) and brain metastases: Preliminary results from the GETUG-AFU 26 (Nivoren) study. J Clin Oncol. 2017;35:4563-4563
123. Kamath SD, Kumthekar PU. Immune Checkpoint Inhibitors for the Treatment of Central Nervous System (CNS) Metastatic Disease. Front Oncol. 2018:8
124. Curran MA, Montalvo W, Yagita H, Allison JP. PD-1 and CTLA-4 combination blockade expands infiltrating T cells and reduces regulatory T and myeloid cells within B16 melanoma tumors. Proc Natl Acad Sci U S A. 2010;107:4275-80
125. Taggart D, Andreou T, Scott KJ, Williams J, Rippaus N, Brownlie RJ. et al. Anti-PD-1/anti-CTLA-4 efficacy in melanoma brain metastases depends on extracranial disease and augmentation of CD8+ T cell trafficking. Proc Natl Acad Sci U S A. 2018;115:E1540-9
126. Choo YW, Kang M, Kim HY, Han J, Kang S, Lee JR. et al. M1 Macrophage-Derived Nanovesicles Potentiate the Anticancer Efficacy of Immune Checkpoint Inhibitors. ACS Nano. 2018;12:8977-93
127. QuantumLeap Healthcare Collaborative. I-SPY 2 TRIAL: Neoadjuvant and Personalized Adaptive Novel Agents to Treat Breast Cancer. <. https://clinicaltrials.gov/ct2/show/NCT01042379>
128. Paolillo M, Serra M, Schinelli S. Integrins in glioblastoma: Still an attractive target? Pharmacol Res. 2016;113:55-61
129. Kurz SC, Wen PY. Quo Vadis—Do Immunotherapies Have a Role in Glioblastoma? Curr Treat Options Neurol. 2018;20:14
130. Sadahiro H, Kang K-D, Gibson JT, Minata M, Yu H, Shi J. et al. Activation of the Receptor Tyrosine Kinase AXL Regulates the Immune Microenvironment in Glioblastoma. Cancer Res. 2018;78:3002-13
131. Lisi L, Laudati E, Navarra P, Dello Russo C. The mTOR kinase inhibitors polarize glioma-activated microglia to express a M1 phenotype. J Neuroinflammation. 2014;11:125
132. Ma DJ, Galanis E, Anderson SK, Schiff D, Kaufmann TJ, Peller PJ. et al. A phase II trial of everolimus, temozolomide, and radiotherapy in patients with newly diagnosed glioblastoma: NCCTG N057K. Neuro Oncol. 2015;17:1261-9
133. Wick W, Gorlia T, Bady P, Platten M, van den Bent MJ, Taphoorn MJB. et al. Phase II Study of Radiotherapy and Temsirolimus versus Radiochemotherapy with Temozolomide in Patients with Newly Diagnosed Glioblastoma without MGMT Promoter Hypermethylation (EORTC 26082). Clin Cancer Res. 2016;22:4797-806
134. Chinot OL, Wick W, Mason W, Henriksson R, Saran F, Nishikawa R. et al. Bevacizumab plus Radiotherapy-Temozolomide for Newly Diagnosed Glioblastoma. N Engl J Med. 2014;370:709-22
135. Lombardi G, De Salvo GL, Brandes AA, Eoli M, Rudà R, Faedi M. et al. Regorafenib compared with lomustine in patients with relapsed glioblastoma (REGOMA): a multicentre, open-label, randomised, controlled, phase 2 trial. Lancet Oncol. 2019;20:110-9
136. Røed Skårderud M, Polk A, Kjeldgaard Vistisen K, Larsen FO, Nielsen DL. Efficacy and safety of regorafenib in the treatment of metastatic colorectal cancer: A systematic review. Cancer Treat Rev. 2018;62:61-73
137. Eggen BJL, Raj D, Hanisch U-K, Boddeke HWGM. Microglial Phenotype and Adaptation. J Neuroimmune Pharmacol. 2013;8:807-23
138. Galanis E, Jaeckle KA, Maurer MJ, Reid JM, Ames MM, Hardwick JS. et al. Phase II Trial of Vorinostat in Recurrent Glioblastoma Multiforme: A North Central Cancer Treatment Group Study. J Clin Oncol. 2009;27:2052-8
139. Qiu Y, Li Z, Copland JA, Mehrling T, Tun HW. Combined alkylation and histone deacetylase inhibition with EDO-S101 has significant therapeutic activity against brain tumors in preclinical models. Oncotarget. 2018:9
140. Bezecny P. Histone deacetylase inhibitors in glioblastoma: pre-clinical and clinical experience. Med Oncol. 2014;31:985
141. Krauze A V, Myrehaug SD, Chang MG, Holdford DJ, Smith S, Shih J. et al. A Phase 2 Study of Concurrent Radiation Therapy, Temozolomide, and the Histone Deacetylase Inhibitor Valproic Acid for Patients With Glioblastoma. Int J Radiat Oncol. 2015;92:986-92
142. Iwamoto FM, Lamborn KR, Kuhn JG, Wen PY, Alfred Yung WK, Gilbert MR. et al. A phase I/II trial of the histone deacetylase inhibitor romidepsin for adults with recurrent malignant glioma: North American Brain Tumor Consortium Study 03-03. Neuro Oncol. 2011;13:509-16
143. Raza M, Prasad P, Gupta P, Kumar N, Sharma T, Rana M. et al. Perspectives on the role of brain cellular players in cancer-associated brain metastasis: translational approach to understand molecular mechanism of tumor progression. Cancer Metastasis Rev. 2018;37:791-804
144. Eroglu Z, Holmen SL, Chen Q, Khushalani NI, Amaravadi R, Thomas R. et al. Melanoma central nervous system metastases: An update to approaches, challenges, and opportunities. Pigment Cell Melanoma Res. 2019;32:458-69
145. Sprowls SA, Arsiwala TA, Bumgarner JR, Shah N, Lateef SS, Kielkowski BN. et al. Improving CNS Delivery to Brain Metastases by Blood-Tumor Barrier Disruption. Trends in Cancer. 2019;5:495-505
Author contact
![]() Corresponding author: Dr. Bozena Kaminska, Laboratory of Molecular Neurobiology, Nencki Institute of Experimental Biology of the Polish Academy, Warsaw, Poland; Tel.: +48 22 5892209; b.kaminskagov.pl
Corresponding author: Dr. Bozena Kaminska, Laboratory of Molecular Neurobiology, Nencki Institute of Experimental Biology of the Polish Academy, Warsaw, Poland; Tel.: +48 22 5892209; b.kaminskagov.pl